
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

3.12. Правые и левые формы деревьев, эдафотипы
Популяции у хвойных видов представляют собой систему с внутренней подразделенностью на левые и правые популяции-изомеры. Они имеют доказанные генетические отличия по гетерозиготности и противоположную адаптацию: левые формы предпочитают прямой свет и слабую конкуренцию, а правые – рассеянный и толерантны к конкуренции. Кроме того, левые формы достоверно лучше растут в сухих условиях, а правые – во влажных (Голиков, 2011, 2013). Ниже показаны побеги сосны с разной закруткой мест крепления хвои, так называемым филлотаксисом – левая и правая формы (рис. 3.20).
В оптимальных условиях рост и частота этих форм одинаковы, но если древостой становится густым, то начинают доминировать правые, а если он более редкий – левые формы. Особенно важно здесь то, что они начинают доминировать почти всегда, когда густота соответствует их предпочтениям, причем даже в несвойственных для них условиях по влажности почвы. Т.е. в редких культурах и во влажных условиях вполне могут расти лучше левые формы, хотя это и не их условия по влажности. В то же время в густых посадках и в сухих условиях вполне могут доминировать правые формы деревьев, которые совсем не любят сухости, но хорошо переносят высокую густоту. Это явление раскрывает механизм гомеостаза популяции, и конкуренция влияет на него сильнее, чем влажность или сухость почвы.

L D
Рис. 3.20. Диссимметрия побегов сосны: левая (L) и правая (D) формы (по Голикову, 2014)
Следует особо отметить, что встречаемость правых и левых форм в исследованиях популяций хвойных пород, начиная от сеянцев и кончая плюсовыми насаждениями, ни разу не снижается до нуля и колеблется строго в пределах соотношения 0,38:0,62 (Голиков, 2014). Это близко к «золотому сечению», в котором проявляют себя универсальные законы Вселенной (Чернов, 2013). Получается, что частота правых и левых форм, постоянно меняясь, выступает как механизм гомеостаза популяций, и этот механизм подчинен универсальным законам Вселенной.
Например, в Псковской обл. А. М. Голиковым были исследованы 28-летние культуры в эдатопе С2 (ель) и С3 (сосна), с густотой посадки 1,0 (редкие) и 4,0 тыс. шт./га (густые культуры) плантационного типа. В густых посадках правые формы превосходили левые по объему ствола на 33 %. В редких посадках, наоборот, левые формы превышали правые по объему ствола на 10–23 %. Нужно заметить, что условия С2 и С3 для левых форм подходят мало, но они почему-то в редких посадках все равно развивались лучше правых. Из этого следует важный вывод: в плантационных культурах с малой густотой левые формы будут продуктивнее правых даже во влажных условиях, для них не свойственных. И это в явном виде стратегия гомеостаза популяции: если появляется обилие света, то левые формы выходят в лидеры даже в нетипичных для них влажных условиях, а правые лучше растут в более густых культурах (Голиков, Бурый, 2008; Голиков, Жигунов, 2012).
На этих же участках культур для анализа хода роста ели было изучено 185 моделей (Голиков, Рогозин, 2013). По росту у этих форм выделили 4 типа развития: быстрое (L+, D+); медленное (L–, D–); ускоренное (L– +, D– +) и замедленное (L+ –, D+ –). Оказалось, что в редких культурах большую часть запасов древесины (59,5 %) накапливают левые формы, а в густых – правые (65,2 %). Примечательно явное доминирование запасов у стабильно быстрорастущих левых (L+) и правых (D+) форм, формирующих особенно крупные стволы; именно они являются центрами накопления запаса, и именно на них должны быть направлены усилия лесоводов и селекционеров (рис. 3.21).
Была также изучена генетическая структура культур сосны в брусничном и черничном типах леса, заложенных по схеме 0,6×1,5 м и выращенных без рубок ухода. При исследовании в 65 лет в черничнике сохранность деревьев оказалась 870, в брусничнике – 550 экз. на 1 га. Популяции имели одинаковый уровень наблюдаемой гетерозиготности (Н0), но анализ изопопуляций обнаружил, что в черничнике между Н0 у правых форм и их ростом связь достоверно отрицательная (r = –0,28 и r = –0,32), а у левых положительная (r = 0,31 и r = 0,34). В брусничнике связи были обратными. Далее оказалось, что лидирующие по росту правые формы, т.е. будущие плюсовые деревья, в желательных для них условиях черничника имели низкую гетерозиготность (Н0 = 0,172 ± 0,016), а в менее комфортном брусничнике более высокую (Н0 = 0,230 ± 0,011). Подобная закономерность обнаружена и для левых форм (Голиков, 2014).
Обнаруженные генетические различия крайне важны, и они объясняют универсальный характер реакций генома популяций культур на изменение ценотических условий. Отметим еще раз, что различия в увлажнении этих эдатопов, между которыми как раз и проходит линия разделения предпочтений для правых и левых форм, не помешала правым формам преобладать в несвойственных им сухих условиях брусничника. Видимо, для ценоза их толерантность к конкуренции оказалась намного важнее, чем снижение их роста в этих, более сухих, условиях.
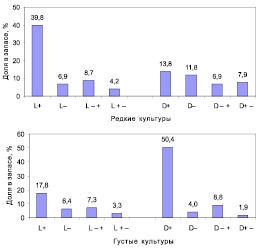
Рис. 3.21. Доли в запасах древесины у правых и левых форм с разным типом их развития в редких и в густых культурах
Показательно в этой связи изучение более тысячи выделов с насаждениями сосны в Среднем Поволжье (Романов, Нуреева, Еремин. 2013), которое показало, что в условиях А2, В2, С2 нормированные запасы в культурах сосны были выше на 12–17 %, с фактическими различиями на 8–49 %. Однако в сухих условиях А1 различия исчезали, а нормированный запас культур оказывался ниже на 4,5 %. Данный факт совершенно непонятен с точки зрения традиционного лесоводства, но объясним с позиций изменения частот левых и правых форм деревьев, и эти позиции следующие.
Правые и левые формы адаптивно неравноценны, и поэтому на сухих почвах чаще встречаются и лучше растут левые, а на влажных почвах правые формы. Но высокая густота культур благоприятствует только правым формам. Поэтому они превосходят левые по встречаемости, хотя условия сухости для них не благоприятны, и они сильно теряют в продуктивности, в особенности на сухих почвах в лишайниковых типах леса.
Эти знания, вместе со знаниями о законах развития древостоев позволяют объединить их в теорию выращивания сосны на сухих почвах со следующими положениями (Рогозин, Голиков, Разин, 2014).
1. При использовании семян, заготовленных в сухих типах леса, в их потомстве мы получаем больше левых форм (на 10–24 %). К 8–10 годам в культурах именно из них формируются деревья-лидеры. В это время кроны смыкаются, что вызывает депрессию их роста и выход в лидеры уже правых форм, толерантных к конкуренции. Если ценоз в этом возрасте разредить и держать деревья в условиях более свободного стояния, то левые формы сохранят своих лидеров и ход роста культур будет самым продуктивным.
2. Если разреживания культур в сухих условиях сделать позже, например, в 20–40 лет, ожидая смыкания крон и «дифференциации» деревьев, как это принято в лесоводстве, то это приведет к доминированию уже правых форм, лидирующих по росту в условиях высокой плотности, но растущих в сухих условиях хуже левых. Левые формы после этого останутся в меньшинстве и древостой понизит продуктивность.
3. При использовании семян, заготовленных в свежих типах леса, можно рассчитывать на получение в потомстве примерно равного соотношения правых и левых форм. Но если древостои будут выбраны густыми, то доля нежелательных для сухих условий правых форм увеличиться.
4. В лесной селекции и семеноводстве необходимо ввести следующее простое правило: семена заготавливают точно в таких же почвенных и ценотических условиях, в каких планируется выращивание лесных культур.
5. В селекции для плантационного выращивания можно рекомендовать отбор плюсовых деревьев в насаждениях, близких по возрасту (50 лет) и по густоте (500–700 шт. /га) к плантационным культурам, с полным совпадением почвенных условий (типа условий местопроизрастания, типа леса).
Таким образом, при исследовании потомства разных эдафотипов, а также плюс-деревьев из разных типов леса, обязательным условием является учет густоты материнского ценоза. При его высокой густоте вполне вероятно получение заурядного потомства. Если же материнский ценоз формировался как разреженное сообщество, то это будет совпадать с условиями выращивания потомства, и его рост будет более быстрым. Положительный эффект будет обусловлен преобладанием в материнской популяции левых форм, для которых плантационные культуры отвечают условиям их наилучшего развития. В оптимальных по увлажнению условиях селекция будет направлена уже на равное соотношение правых и левых форм, а при переувлажнении – на преобладание правых форм (Голиков, 2014; Рогозин, Голиков, Разин, 2014).