
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

4.2. Наследуемость у сосны в разном возрасте
В исследованиях потомства сосны постоянных семенных участков (ПЛСУ) тип связи между высотой матери и ее потомством оставался неясен даже на выборках из 120 пар данных, а близкие к нулю корреляции не давали возможности выяснить ее тренд. Ниже мы приводим некоторые данные, которые помогут читателю получить общее представление о коллизиях наследуемости у сосны.
Первые массовые исследования для 387 материнских деревьев сосны в 3- и 4-летнем возрасте их потомства показали, что корреляции высот мать-семья составили: для Пермского ПЛСУ r1 = 0,12 ± 0,07 и для Левшинского участка r2 = 0,16 ± 0,076. Коэффициенты регрессии были равны b1 = 0,14 ± 0,07 и b2 = 0,105 ± 0,06. Усреднение значений b и r дает оценки наследуемости в узком смысле h2 = 0,26 ± 0,14.
В опытах с испытаниями потомства двух других ПЛСУ были получены уже другие значения r, включая и отрицательные. Рассмотрим вначале корреляции мать-семья в потомствах Очерского ПЛСУ от двух урожаев, в тест-культурах на супеси, суглинке и песчаных почвах (рис. 4.1).
Сразу отметим, что корреляции на двух верхних полях в среднем несколько выше, и пока возьмем на заметку, что условия тест-культур там (Б2 и С2) оказались близки по увлажнению к условиям на лесосеменном участке (С2), в сравнении с нижними графиками, где испытания проводили в более сухих условиях А1, и где одна из корреляций опустилась до нуля.
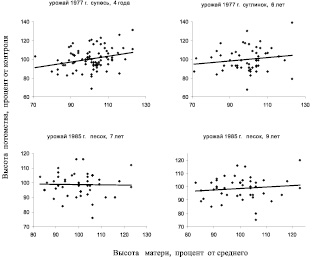
Рис. 4.1. Наследуемость в потомстве деревьев сосны Очерского ПЛСУ
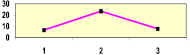
Далее был поставлен простой вопрос и далее решен в виде статистической игры – как будут меняться значения r и линии тренда, если выборки уменьшить и искусственно разделить на две равные части? Для этой игры использовали Нижне-Курьинское потомство урожая 1985 и 1982 г. при возрасте семей 7, 9 и 18 лет (рис. 4.2).
Отметим, что в средней паре графиков связь из отрицательной (–0,09) становится положительной (0,19) и эти колебания корреляций укладываются в доверительный интервал их значений при оценке по общей выборке, из которой они взяты. Поэтому обнаруживаемые в каждом конкретном потомстве возрастания или падения r с возрастом или на другой почве, при недостоверных значениях этой связи, точно также недостоверны и отражают ее случайные колебания. Наблюдаемые изменения наклона линий тренда, вплоть до смены знака корреляций, дают основания сомневаться в наличии «наследуемости» в ряде выборок, которая для них оказывается недостоверной и поэтому недоказанной.
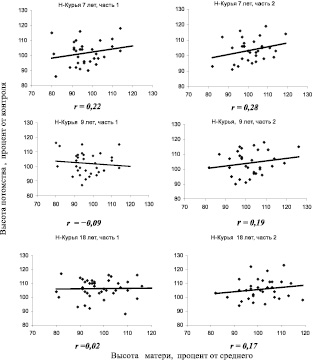
Рис. 4.2. Статистическая игра – две выборки из одной совокупности (части 1 и 2) в Нижне-Курьинском потомстве в испытательных культурах в разном возрасте; внизу приведены коэффициенты корреляции
Но, с другой стороны, у нас имеется 13 выборок, и их можно объединить, увеличив объем выборок, и далее доказать наличие или отсутствие связей в целом, уже на большой выборке. Однако возраст семей по опытам был разным, поэтому вначале поместим значения корреляций на обычную точечную диаграмму, с целью разобраться в этом вопросе. Оказалось, что есть видимая тенденция падения этих корреляций (рис. 4.3).
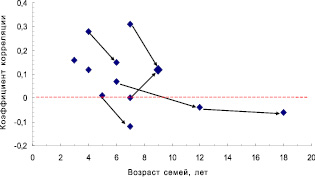
Рис. 4.3. Снижение корреляций высот мать-семья с возрастом в 13 выборках; стрелки соединяют повторные значения в более позднем возрасте
На этом рисунке видно, что при повторных измерениях семей в более позднем возрасте корреляции снижаются в 5 случаях из 6; еще в одном случае, на родственных потомствах Пермского и Левшинского ПЛСУ, заложенных семенами одного происхождения, падение r также имеет место (две точки, не соединенные стрелкой).
Данные по этим корреляциям можно разбить для анализа на две группы: для раннего возраста 3–6 лет и для возраста 7–18 лет (табл. 4.1).
В раннем возрасте, в 3–6 лет, средневзвешенная корреляция между высотами матерей и их семей по 727 парам данных оказалась равна r = 0,125 ± 0,036 и достоверна (t = 3,6 > t0,99 = 2,63). Однако далее, в 7–18 лет, среднее по 491 паре данных значение r = 0,020 ± 0,045, т.е. корреляция оказалась практически нулевой и недостоверной (t = 0,44 < t0,90 = 1,66).
При сравнении этих двух корреляций, полученных для разного возраста, получаем, тем не менее, достоверное различие между ними t = 1,84 > t0,90 = 1,66. Поэтому видимое на рис. 4.3 падение корреляций мать-семья с возрастом также оказывается достоверным, несмотря на одно из довольно высоких значений r = 0,31 в 7-летнем возрасте.
Таблица 4.1
Корреляции высот мать-семья в раннем (3–6 лет) и более старшем (7–18 лет) возрасте у сосны обыкновенной
|
Возраст семей 3–6 лет |
Возраст семей 7–18 лет |
||||
|
Возраст, лет |
Число семей, шт. |
r |
Возраст, лет |
Число семей, шт. |
r |
|
3* |
171 |
0,16 |
7 |
45 |
0 |
|
4 |
84 |
0,28 |
7* |
133 |
|
|
4* |
216 |
0,12 |
7 |
64 |
0,31 |
|
5* |
133 |
0,01 |
9 |
45 |
0,12 |
|
6 |
53 |
0,15 |
9 |
64 |
0,12 |
|
6 |
70 |
0,07 |
12 |
70 |
|
|
18 |
70 |
||||
|
Итого пар данных |
727 |
491 |
|||
|
Значение r средневзвешенное |
0,125 |
0,020 |
|||
|
Ошибка r |
0,036 |
0,045 |
|||
|
Достоверность, t |
3,6 |
0,44 |
|||
Примечание. * – испытания проведены в школе питомника.
Как видим, уже начало анализа убеждает нас, насколько изменчива связь мать-потомство и как случайные факторы меняют ее знак, да и вообще само ее наличие. При этом о «наследуемости» можно говорить, по-видимому, только как о материнском эффекте, который проявляется лишь в первые годы.
Особо отметим, что результаты эти получены в случайных, а не «плюсовых» выборках. Если их сравнивать с результатами, полученными другими авторами на выборках с преобладанием плюс-деревьев, то становятся понятны причины разных мнений лесных селекционеров в отношении того, принимать или не принимать идею плюсовой селекции в качестве главного направления работ. Всегда жаль затраченных усилий, и тяжело расставаться с иллюзиями. Дело в том, что в период развертывания работ по лесной селекции в России в 1980-е гг. обычно выбирали 20–30 плюсовых деревьев и контроль в виде семян производственного сбора. В таком опыте наследуемость не определяли, так как выборка была усечена со стороны обычных деревьев с малыми и средними размерами (неслучайная выборка).
Так, в серии из 5 опытов (Ефимов, 1997) в качестве примера рассматривался один из них, с представленностью потомства от 19 плюс-деревьев. В этом опыте среди 18 последовательных корреляций мать-семья в возрасте до 20 лет только в 1 случае корреляция достигла достоверного уровня (r = 0,63). Однако по этим данным, тем не менее, был сделан вывод «…о заметном аддитивном вкладе генов в изменчивость потомств по высоте», и о том, что «они свидетельствуют об определенной эффективности массового отбора сосны на быстроту роста, но этот эффект проявляется у потомств в возрасте, близком к 1/3 возраста рубки. Предварительную генетическую оценку плюсовых деревьев сосны по росту полусибовых семей можно проводить не ранее 20-летнего возраста» (Ефимов, 1997).
На эти выводы ориентируется и сейчас множество исследователей, и в них проскальзывает что-то близкое к запрету даже на саму мысль о возможности правильных генетических оценок матерей по их потомству в возрасте менее 20 лет. Однако выводы эти были основаны на малых выборках, где корреляции оказались недостоверны. В результате и запрет, и сами рекомендации в итоге получились совершенно неубедительными.
Так как рассмотренные выше корреляции меняли тренд при статистической игре (см. рис. 4.2), а также при очистке выборки всего лишь от 1–2 уклоняющихся значений, то непростой характер наследуемости помог раскрыть детальный анализ данных на самих полях корреляции. Анализ этот известен как «анализ долей» и применяется для анализа качественных признаков. У него есть ценные качества – он чувствительнее, проще и понятнее, чем более сложный дисперсионный анализ.
Отметим, что на всех ПЛСУ изучали деревья без предпочтений какому-либо фенотипу; к тому же неожиданно оказалось, что, несмотря на создание семенных участков лучшими сеянцами, а далее разреживание с оставлением лучших деревьев, высоты деревьев на них в 24–26 лет менялись в пределах от 72 до 124 % от среднего значения. Такой большой размах высот у сосны оказался близок к колебаниям ее высот в обычных древостоях. Поэтому полученные выше корреляции можно оценивать как репрезентативные для потомства сосны при возрасте матерей 24–26 лет.
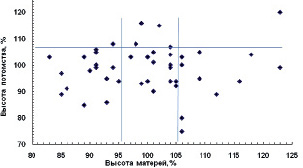
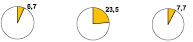
Отмеченный размах высот у матерей на ПЛСУ оказался во многом неожиданным, так как в условиях свободного стояния можно было ожидать формирование более выравненных деревьев. Такой их размах, однако, позволил далее хорошо разделить деревья на градации их высоты: низкие, средние, высокие и самые высокие. Для примера приведем потомство, где явное преимущество показали средние матери: лучших семей у них оказалось в 3 раза больше (23,5 %), чем в соседних градациях, где их частота составила 6,7 % у низких и 7,7 % – у высоких матерей (рис. 4.4).
а
б
в
Рис. 4.4. Высота матерей в 24 года на Очерском ПЛСУ и высота их потомства в 9 лет (а), доли (б) и частоты (в) лучших семей в % в градациях высот матерей: 1 – низкие, 2 – средние, 3 – высокие матери
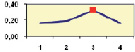
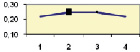
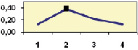
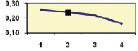
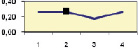
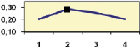
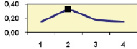
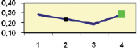
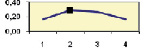
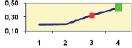

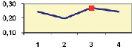
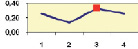
Рис. 4.5. Частота лучших семей (в долях) в потомствах сосны ПЛСУ: а, б – Очерский, урожай 1977 г. в 4 и 6 лет; в, г – Очерский, урожай 1985 г.
в 7 и 9 лет; д – Левшинский в 3 года; е – Пермский в 4 года; ж, з, и – Нижне-Курьинский, урожай 1982 г. в 6, 12 и 18 лет; к, л – то же потомство в 5 и 7 лет в школе; м, н – Нижне-Курьинский, урожай 1985 г. в 7 и 9 лет в школе. По горизонтальной оси обозначены цифрами градации высоты матерей: 1 – низкие; 2 – средние; 3 – «просто высокие»; 4 – самые высокие
Столь большой всплеск числа лучших семей у средних матерей был не случаен, и таких случаев оказалось много. В более чем половине испытаний, которые мы рассмотрим далее, именно средние по высоте матери давали повышенное число лучших семей.
Ниже показаны аналогично рассчитанные частоты лучших семей на других полях корреляций в 13 опытах с 45–216 парами данных в каждом, где было изучено 604 потомства сосны четырех семенных участков. Для приведенных частот использовано их усреднение при двух интенсивностях отбора лучших семей: 1/3 и 1/7, или 33 и 15 % (рис. 4.5).
Из этих 13 лишь на 5 графиках доля лучших семей повышена у «просто высоких» и самых высоких матерей (38 % случаев). В большинстве же случаев (8 графиков или 62 %) эта доля выше у средних матерей. Вместе с тем, и это очень важно для реабилитации идеи плюсовой селекции, низкие по высоте матери практически в половине случаев (7 из 13) давали пониженную долю лучших семей (см. рис. 4.5).
Если же рассмотреть частоты лучших семей по двум группам, в раннем возрасте (3–6 лет) и в старшем (7–18 лет), то результаты становятся еще определеннее для понимания тенденций наследуемости.
В раннем возрасте как низкие, так и самые высокие матери дают пониженную частоту лучших семей (75 %), тогда как средние и «просто высокие» ее увеличивают до 109–115 % (табл. 4.2).
Таблица 4.2
Частота лучших семей сосны у матерей с разной высотой (выделены достоверные превышения)
|
Возраст, лет |
Число семей, шт. |
Корреляция высот мать-семья (r) |
Частота лучших семей у матерей |
|||
|
низкие |
средние |
«просто высокие» |
самые |
|||
|
6 |
53 |
0,15 |
53 |
159 |
89 |
53 |
|
4 |
84 |
0,28 |
68 |
79 |
137 |
68 |
|
3* |
171 |
0,16 |
68 |
122 |
110 |
68 |
|
4* |
216 |
0,12 |
72 |
120 |
106 |
72 |
|
6 |
70 |
0,07 |
106 |
55 |
138 |
106 |
|
5* |
133 |
0,01 |
85 |
120 |
107 |
85 |
|
Итого |
727 |
|||||
|
В среднем |
0,125** |
75 |
109 |
115 |
75 |
|
Примечания: * – испытания в школе питомника; ** – средневзвешенное значение.
С увеличением возраста потомства, когда корреляции мать-семья понижаются до близких к нулю значений (см. рис. 4.1), частоты лучших семей практически выравниваются и имеют значения 96–106 % (табл. 4.3).
Таблица 4.3
Частота лучших семей сосны в возрасте 7–18 лет у матерей разной высоты (достоверные превышения выделены)
|
Возраст, лет |
Число семей, шт. |
Частота лучших семей у матерей разной высоты, процент от нормы |
||||
|
низкие |
средние |
«просто |
самые |
|||
|
9 |
45 |
0,12 |
63 |
139 |
73 |
63 |
|
7 |
45 |
0 |
111 |
112 |
73 |
111 |
|
12 |
70 |
–0,04 |
92 |
104 |
104 |
92 |
|
18 |
70 |
–0,06 |
108 |
100 |
92 |
68 |
|
7* |
133 |
–0,12 |
120 |
100 |
80 |
120 |
|
7 |
64 |
0,31 |
80 |
82 |
136 |
182 |
|
9 |
64 |
0,12 |
103 |
83 |
114 |
103 |
|
Итого |
491 |
|||||
|
В среднем |
0,02** |
97 |
103 |
96 |
106 |
|
Примечания: * – испытания в школе питомника; ** – средневзвешенное значение.
Рассмотрение данных можно завершить общей таблицей, которая оказалась довольно сложной для таких разнообразных опытов (табл. 4.4).
Ее итоги показывают, что самые высокие матери уменьшают частоту лучших семей до 92 % от среднего, и это чуть больше, чем у низких матерей (87 %). В то же время средние и «просто высокие» матери ее повышают до 105–106 %. Это немного, но тенденция ясна – это стабилизирующий отбор.
Заключение о наследуемости у сосны после столь сложного анализа экспериментальных данных будет следующим. Корреляции высот мать-семья у сосны в возрасте потомства до 18 лет при возрасте матерей 24–26 лет слабые и недостоверные и есть явные признаки того, что связи криволинейны. Поэтому для оценки наследуемости коэффициент корреляции непригоден. Более правильной будет оценка по доле лучших семей, наблюдаемых у матерей с разными высотами: низких, средних, «просто высоких» и «самых высоких» (в какой-то мере «плюсовых»). В целом полученные результаты особенно выразительно показывают нецелесообразность стремления к отбору самых высоких матерей – они дают лучшие семьи лишь в 38 % случаев; в остальных 62 % случаев преимущество оказывается у средних и «просто высоких» матерей. Вместе с тем низкие матери в большинстве случаев давали пониженную долю лучших семей. Это подводит нас к мысли о том, что у сосны обыкновенной «плюсовая» селекция (отбор самых высоких родителей) не является единственно лучшим вариантом ее селекции, по крайней мере, в изученных ценопопуляциях постоянных лесосеменных участков.
Таблица 4.4
Оценка наследуемости высоты у сосны по коэффициенту корреляции (r) и частоте лучших семей в градациях высот матерей
|
ПЛСУ, тип условий |
Год урожая |
Выращивание сеянцев |
Число вариантов |
Тест-культуры |
r |
Частоты лучших семей у матерей разной высоты, % от нормы |
||||
|
ТУМ |
воз |
низкие |
средние |
«просто |
самые |
|||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
|
Очерский, ТУМ С2 |
1977 |
тепл 2 |
84 |
Б2 |
4 |
0,28 |
68 |
79 |
137 |
68 |
|
пит 2 |
53 |
С2 |
6 |
0,15 |
53 |
159 |
89 |
53 |
||
|
1985 |
пит 2 |
45 |
А1 |
7 |
0,00 |
111 |
112 |
73 |
111 |
|
|
9 |
0,12 |
63 |
139 |
73 |
63 |
|||||
|
Левшинский ТУМ С2 |
1979 |
пит 1 + шк 2 |
171 |
Б2 |
3* |
0,16 |
68 |
122 |
110 |
68 |
|
Пермский, ТУМ С2 |
1981 |
пит 2 + шк 2 |
216 |
Б2 |
4* |
0,12 |
72 |
120 |
106 |
72 |
|
Нижне-Курьинский, ТУМ Б2 |
1982 |
пит 1 + шк 2 |
70 |
Б2 |
6 |
0,07 |
106 |
55 |
138 |
106 |
|
12 |
–0,04 |
92 |
104 |
104 |
92 |
|||||
|
18 |
–0,06 |
108 |
100 |
92 |
68 |
|||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
|
Нижне-Курьинский, ТУМ Б2 |
1982 |
тепл 1 |
133 |
Б1 |
5* |
0,01 |
85 |
120 |
107 |
85 |
|
7* |
–0,12 |
120 |
100 |
80 |
120 |
|||||
|
1985 |
пит 2 |
А1 |
7 |
0,31 |
80 |
82 |
136 |
182 |
||
|
9 |
0,12 |
103 |
83 |
114 |
103 |
|||||
|
В среднем по 13 опытам, включая измерения в разном возрасте |
8,7 |
0,08 |
87 |
106 |
105 |
92 |
||||
|
Всего вариантов/семей |
836/604 |
|||||||||
Примечания: пит 1 – питомник 1 год; шк 2 – школа 2 года; *– выращивание в школе; ** – матери с высоким выходом семян из шишек; выделены достоверные превышения.