
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

4.5. Наследуемость у ели из естественных популяций
В естественных насаждениях, где были изучены 7 популяций и 300 плюс-деревьев, коэффициент наследуемости высоты нам рассчитать не удалось, так как, в связи с большим объемом работ по заготовке образцов семян, контрольные деревья в них не выделялись. Но какое-то влияние размеров матерей на рост потомства здесь все-таки можно выяснить, с оговорками о том, что их размеры определял, конечно же, их возраст, который колебался от 65 до 150 лет, а также в меньшей степени условия местопроизрастания, классифицированные как С2 (ельники кисличники) и С3 (ельники липняковые). Влияние возраста было тем более интересно, что, несмотря на примерно одинаковые условия, высоты плюс-деревьев колебались от 25 до 36 м.
В тест-культурах плантационного типа, а также в обычных культурах («редких» и «густых») корреляции высот мать-семья оказалась близки к нулевым значениям и недостоверны (0,020 и –0,012 соответственно). Коэффициент наследуемости по этим значениям рассчитывать некорректно, так как его значение будет смещено из-за отсутствия в выборке материнских деревьев средних и малых размеров. Однако «нулевые» корреляции все-таки показывают нам, что при увеличении высот матерей высоты их семей не увеличиваются. Не удовлетворившись корреляционным анализом, можно использовать далее анализ долей. Размах высот у плюсовых деревьев составлял 11 м, и целесообразно разделить их на три градации по высоте (табл. 4.7).
Таблица 4.7
Частота лучших семей в потомстве плюс-деревьев ели, отобранных в естественных насаждениях в возрасте 65–150 лет
|
Показатели |
Градации высот плюсовых деревьев |
Вся выборка |
||
|
средние |
просто высокие |
самые |
||
|
Пределы высот у матерей, м |
25–28,5 |
29–31 |
31,5–36 |
25–36 |
|
Потомство в тест-культурах плантационного типа (редкие культуры) |
||||
|
Всего семей в градации, шт. |
82 |
131 |
87 |
300 |
|
В т.ч. с высотой 115 % и более, % |
8,5 |
7,6 |
6,9 |
7,67 |
|
Отношение к средней частоте |
1,11 |
1,00 |
0,90 |
1,00 |
|
Потомство в тест-культурах обычного типа (густые культуры) |
||||
|
Всего семей в градации, шт. |
69 |
105 |
71 |
245 |
|
В т.ч. с высотой 115 % и более, % |
27,5 |
18,1 |
18,3 |
20,8 |
|
Отношение к средней частоте |
1,32 |
0,87 |
0,88 |
1,00 |
И здесь неожиданно средние по размерам плюс-деревья (т.е. самые низкие в этой выборке), в сравнении с просто высокими и самыми высокими, дали повышенное число лучших семей: в редких культурах 8,5 % против 6,9–7,6 %, а в густых культурах 27,5 % против 18,1 и 18,3 % у высоких и самых высоких. Это в 1,11–1,32 раза больше, чем в целом по всей выборке. Различия между градациями еще больше и достигают в первой выборке 1,2 раза, а во второй – 1,5 раза, и различия этих частот достоверны. Сразу отметим, что эти данные подтверждают нецелесообразность доктрины обязательного стремления к отбору самых высоких родителей; достаточен отбор просто высоких матерей, о чем уже говорилось выше как для ели из популяций культур Теплоуховых, так и для сосны.
Как же объяснить эти падения частот? Попытки разделить общую выборку на выборки по ценопопуляциям (происхождениям), которых насчитывалось 7, не дали результата в связи с сокращением числа наблюдений в градациях и, как следствие, появлением резких колебаний в долях лучших семей.
Вместе с тем была надежда, что если снять (обнулить) влияние возраста матерей и перейти к среднему приросту материнского дерева, то это должно внести некоторую ясность по вопросу влияния скорости роста матерей на рост их потомства. Согласно основной идее плюсовой селекции, чем выше материнское дерево (и средний прирост дерева-матери), тем быстрее должно расти и потомство. Однако переход на прирост дал еще более неожиданные результаты (рис. 4.10).
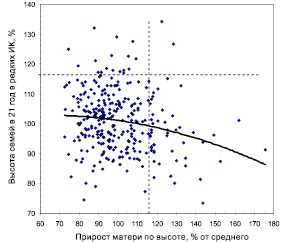
Рис. 4.10. Прирост по высоте плюс-деревьев ели из естественных популяций и высота их потомства в тест-культурах плантационного типа
(отрицательная наследуемость скорости роста в высоту)
Показанная на этом рисунке корреляция между приростом матери по высоте и высотой семей оказалась r = –0,185 ± 0,056. Связь отрицательна и достоверна при t = 3,32 > t0,05 = 2,0. Однако на участке густых тест-культур она оказалась практически нулевой (r = 0,03).
Далее у этих матерей был расчитан и прирост по диаметру, и его влияние на высоту семей было еще отрицательнее (r = –0,209 ± 0,055), а в густых культурах оно также было близким к нулю (r = 0,06).
Объяснить этот весьма неожиданный экспериментальный факт, тем не менее, можно, если полагать, что потомствам некоторых плюс-деревьев (восемь точек в правой части поля) условия в культурах плантационного типа «не нравятся» и подходят мало, и их потомство снижает там высоту.
Все эти явления наталкивают на мысль о том, что здесь «работает» эпигенетика или, как мы образно пояснили выше, проявляет себя в потомстве некая «память» о том, в каких условиях развивались их родители. По-видимому, для ели, происходящей из естественных насаждений, больше подходят густые культуры и ее наследуемость в них нейтральна, а в более редких плантационных посадках она меняет свой рост, и ее наследуемость становится отрицательной. Поэтому ель из естественных насаждений менее подходит для программы селекции с ориентацией на плантационное выращивание; здесь лучше использовать исходный материал из старых культур, где «память» потомства уже настроилась на выращивание в более комфортных условиях ослабленной конкуренции, и в разреженных культурах плантационного типа ее наследуемость оказалась на вполне приличном уровне, с корреляцией высот мать×семья r = 0,185 ± 0,065, о чем шел разговор выше.
В следующих разделах мы покажем, во-первых, что образ «памяти» взят не случайно, и он связан, скорее всего, с перераспределением в родительских древостоях частоты правых и левых форм деревьев, у которых генетически противоположная адаптация к конкуренции: левые предпочитают ее в ослабленном виде, а правые к ней вполне толерантны.
Во-вторых, мы уже приводили данные о том, что ель из культур Теплоуховых имеет больше признаков ели европейской, и поэтому она тяготеет к эволюции по другому типу, чем ель из естественных насаждений, имеющая шишки и семена меньших размеров, что характерно для ели сибирской. Заметим, что по лесосеменному районированию 1982 г. ель в Пермском крае была отнесена к ели сибирской. Поэтому для этого вида ели, а также для сосны обыкновенной отрицательная наследуемость просто есть, и этот экспериментальный факт следует учитывать в генетике и селекции этих хвойных видов, имеющих самое широкое промышленное использование и ареал на территории России.