
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

6.4. Плотность подроста ели на пробной площади кв. 30
В табл. 6.4 приведены исходные данные для моделирования.
|
Таблица 6.4
Плотность подроста ели
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Снова, как это было в предыдущих примерах, минимальная средневзвешенная плотность подроста наблюдается около материнской ели, то есть в центре круговой пробной площади с материнской елью.
Но максимум плотности размещения, то есть верхняя граница доверительного интервала, сместился далеко от центра, вместо 20 м в три раза дальше на 60 м.
Север. По данным табл. 6.4 и графику на рис. 6.14 в соответствии с биотехнической закономерностью вида
![]()
![]() (6.15)
(6.15)
два стрессовых возбуждения стали ближе и дальше от материнской ели и получили по одинаковому максимуму 7,5 шт./ар на 20 и 50 м.
Однако ближнее возбуждение является более резким из-за двукратного увеличения половины периода волны у дальнего стресса.
По остальным сторонам света происходило заметное нарушение биоэнергетических связей.
|
Рис. 6.14. График изменения плотности подроста ели на пробной площадке в зависимости от расстояния в северном направлении |
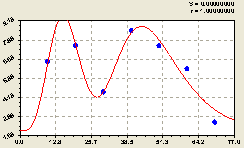
Северное направление примечательно еще и тем, что соблюдается закон сохранения энергии. Дело в том, что резкие возмущения с двумя пиками привели к значительному спаду плотности размещения подроста материнской елью на расстоянии 70 м. По сравнению с другими сторонами света результативность генерации отпрысков снизилась по северному направлению в 2-3 раза.
Восток. По рис. 6.15 и биотехнической закономерности вида
![]()
![]() (6.16)
(6.16)
|
Рис. 6.15. График изменения плотности малых деревьев ели на пробной площадке в зависимости от расстояния в восточном направлении |
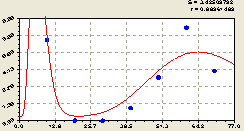
заметно, что амплитуда процесса генерации себе подобных у материнской ели на восточном направлении значительно повысилась. В механике такое колебание носит название «идти в разнос». В итоге появились места с нулевой плотность подроста ели. Но пока график еще не переходит через ось абсцисс в отрицательную полуплоскость.
Юг. Из рис. 6.16 и формулы вида
![]() (6.17)
(6.17)
заметно, что, строго говоря, оба стресса превратились в волновые возмущения. И это видно по расстановке точек вокруг графика на рис. 6.16.
Такое размещение показывает, что нужно переходить к идентификации вейвлет-сигналов, то есть к волновым уравнениям с четко выраженными циклами и амплитудно-частотными характеристиками.
Вначале мы не предполагали излагать в данной книге волновую теорию роста и развития лесных деревьев внутри лесного древостоя, причем с образованием внутри себя отдельных микроскопических сообществ в идее биогрупп из деревьев разных пород и одной породы особей.
|
Рис. 6.16. График изменения плотности малых деревьев ели на пробной площадке в зависимости от расстояния в южном направлении |
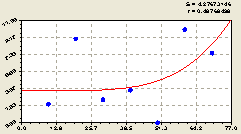
Причем заметно из расположения точек около графика на рис. 6.16, что на выделе 9 кв. 30, из-за значимого антропогенного влияния, трудно будет выявлять вейвлет-сигналы из-за хаотического распределения плотности деревьев.
Запад. Поэтому, на что указывают рис. 6.17 и биотехническая закономерность по формуле вида
![]()
![]() , (6.18)
, (6.18)
|
Рис. 6.17. График изменения плотности малых деревьев ели на пробной площадке в зависимости от расстояния в западном направлении |
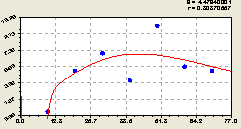
на более экологически структурированном древостое вполне возможна волновая теория.
Это мы покажем полностью на параметрах круговой пробной площади выдела 8 лесного квартал 37.
Но главный методологический вывод можно сделать и из графика на рис. 6.17. Это - принцип соразмерности структуры и параметров популяции с её частью в виде подроста.
Поэтому известная проблема «отцов и детей», а в ельнике точнее проблема «матерей и их детей» вполне решаема измерениями абсолютной численности (при постоянной площади прямоугольных пробных площадок) или же плотности (при переменой площади различных форм и размеров у биогрупп лесных деревьев).
Всего на кольцах. Этот вывод подтверждается на рис. 6.18 и в конструкции биотехнической закономерности в виде формулы
![]()
![]() . (6.19)
. (6.19)
|
Рис. 6.18. График изменения плотности подроста ели на кольцевой пробной площадке в зависимости от расстояния |
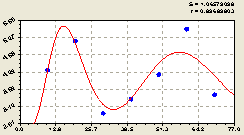
Здесь также заметно, что подрост по густоте размещения получил аналогичную для всей популяции ели биотехническую закономерность.
Этот факт указывает на то, что возможны три способа измерений: всех деревьев, взрослых деревьев ели или же только подроста ели. Все они должны по параметрам распределяться на лесном участке по одним и тем же формулам, но с разными значениями их коэффициентов (параметров модели). Но как быть с искусственными посадками, где человек чрезмерно усложнил биотехнические закономерности, мы пока не знаем.
Пространственное распределение на рис. 6.19 показывает расположение биогрупп подроста ели по некоему кольцу.
|
Рис. 6.19. Изменение плотности подроста ели на пробной площади от расстояния и азимута |
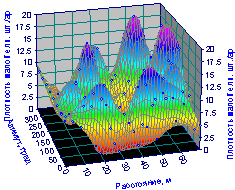
Назовем кольцо из восьми заметных групп малой ели условно как «кольцо генерации».
Заметно, что по сравнению с рис. 6.13, центральный малый выступ в этом кольце генерации пропал. Но зато по краям кольца четко проявились еще два сгущения подроста.
Может оказаться, что фактически материнская ель, то есть семенное дерево, когда-то давно, находилось именно в центре этого кольца генерации.
Но для доказательства этой гипотезы нужны дополнительные измерения, причем с учетом как свежих и спелых пней, а также мест на территории круговой пробной площади с заметными признаками нахождения в этой точке сгнившего пня материнской ели.
Таким образом, круг предметов исследования расширяется.