
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

1.1. Значение околоцветника
Околоцветник выполняет функции привлечения насекомых и защиты генеративных органов. Сторонники стробильной теории придают решающее значение энтомофилии в возникновении цветка покрытосеменных. С возникновением энтомофилии к первичной защитной функции околоцветника добавляется функция привлечения насекомых (Arber, Parkin, 1907). По мнению А.Л. Тахтаджян (1954) эволюция покрытосеменных связана с насекомыми, а цветок возник из энтомофильного стробила (шишки) и является продуктом энтомофилии. Возникновение специализированных насекомых-опылителей привело к прогрессу в перекрестном опылении - к усилению постоянства посещения насекомыми цветков одного и того же вида растений при каждом вылете. В то же время развитие постоянства посещения цветков насекомыми способствовало прогрессу в их строении. Наиболее высокое постоянство посещения устанавливается у цветков со сростнолепестным венчиком, у которых уменьшается число тычинок и увеличиваются возможности для выделения нектара и его укрытия у основания трубки венчика. Оно осуществляется пчелами, длиннохоботными мухами и бабочками. Постоянство посещения имеет преимущество как для цветков, которые лучше опыляются и меньше расходуют пыльцы, так и для насекомых, находящих здесь корм.
С взаимным приспособлением растений и насекомых сопряжено образование «шпор» на нижней губе венчика, которые являются резервуарами для нектара (Jones, 1950).
Яркая окраска частей околоцветника обусловлена не только в связи с опылением насекомыми, так как цветки многих ветро-опыляемых растений также яркоокрашены. Например, в шишках ели, сосны и лиственницы содержится большое количество антоцианов. Встречаются они в генеративных органах ольхи, осина, ясеня (Н.В. Первухина, 1979).
По мнению В. Г. Александрова и М. И. Савченко (1951), возникновение антоцианов, в первую очередь, нужно связывать не с энтомофилией, а с особенностями физиологии цветка, так как чашелистики и лепестки участвуют в физиологических процессах, протекающих при формировании половых клеток. И. В. Мичурин (1948) указывал, что удаление околоцветника отрицательно сказывается на энергии прорастания пыльцы и завязывании плодов. Например, у крыжовника прорастание пыльцы при кастрации с оставлением околоцветника наблюдалось через 2 ч. после опыления, тогда как пыльца, нанесенная на рыльца пестиков цветков, кастрированных с удалением околоцветника, начала прорастать через 8 ч.
По данным Е.Н. Соколовой (1953), в опытах с грушей завязывание плодов при кастрации с оставлением околоцветника составляло 50,5% по числу завязавшихся и 8,4% по числу сформировавшихся плодов, тогда как при кастрации с удалением околоцветника число завязавшихся плодов насчитывало 45%, а сформировавшихся - 6,25%. Следовательно, с оставлением околоцветника число завязавшихся плодов было на 5,5%, а сформировавшихся на 2,15% больше, чем с удалением околоцветника. Таким образом, удаление околоцветника отрицательно сказывается на прорастании пыльцы, препятствуя опылению и формированию плодов.
Согласно имеющимся литературным данным, негативное влияние удаления лепестков связано с нарушением физиологической системы цветка, оказывающим существенное влияние на формирование половых клеток. И. Н. Голубинский (1950), считает, что летучие выделения околоцветника оказывают положительное влияние на прорастание пыльцевых зерен. Проращивая пыльцу в чашках Петри, с помещенными в них лепестками, он установил, что процент проросших пыльцевых зерен и длина пыльцевых трубок были значительно большими в чашках Петри с помещенными в них лепестками, по сравнению с контрольными чашками, в которых лепестки отсутствовали. Так, число проросших пыльцевых зерен под влиянием выделений лепестков, помещенных в чашки, увеличилось на 23,2% у смородины черной и на 34,6% - у бузины черной. Длина пыльцевых трубок, соответственно возросла с 158 до 215 и с 318 до 502 мкм. Стимулирующее влияние летучих выделений околоцветника наблюдалось и в другом варианте опыта, когда открытые чашки Петри, с проращиваемой в них пыльцой, помещались в крону цветущего растения. Например, у дикой яблони число проросших пыльцевых зерен увеличилось на 28,9% по сравнению с контрольными чашками, находившимися на открытом воздухе, длина пыльцевых трубок возросла почти втрое.
Другой физиологической особенностью околоцветника является динамика крахмала в лепестках. Некоторые исследователи указывают на обилие крахмала и глюкозы в лепестках бутонов многих растений и почти полное отсутствие этих веществ в околоцветнике распустившегося цветка. Так, Muller, (1893), считает, что лепестки бутонов магнолии содержат значительное количество крахмала и глюкозы. По данным А.В. Добротворской (1962), клетки лодикул в неоплодотворенном цветке пшеницы имеют высокое содержание крахмала. После оплодотворения крахмал в лодикулах исчезает. Клетки околоцветника бутона гладиолуса также содержат большое количество крахмала (рис. 1), который почти полностью отсутствует в околоцветнике распустившегося цветка (рис. 2).
Кроме крахмальных зерен околоцветник содержит большое количество зеленых пластид. При этом в чашелистиках хлоропласты находятся на протяжении цикла развития цветка, а в лепестках - в период бутонизации. В лепестках хлоропласты имеют бледнозеленую окраску и содержат большое количество крахмала. По мнению В.Г. Александрова и др. (1947) они способствуют гидролитическим реакциям, содействуя перемещению углеводов по тканям цветка. Зеленые пластиды чашелистиков отличаются от пластид лепестков по интенсивности окраски и продолжительности существования. Исследование Н.В. Первухиной (1979) хлоропластов цветковых чешуй и листа пшеницы показало, что пластиды цветковых чешуй имеют такое же строение, как и пластиды листа. Они окружены двойной оболочкой, содержат межгранные ламеллы и граны, которые по числу тилакоидов не отличаются от гран хлоропластов листа. В матриксе хлоропластов цветковых чешуй имеются рибосомы, встречаются осмиофильные глобулы. Сходство субмикроскопической морфологии пластид чешуи и листа свидетельствует об их функциональной близости.
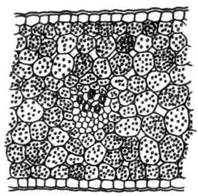
Рис. 1. Участок поперечного среза доли околоцветника Gladiolus в период бутонизации (в клетках содержится крахмал) (по Н.В. Первухиной, 1979)
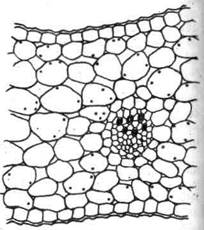
Рис. 2. Участок поперечного среза доли околоцветника раскрывшегося цветка Gladiolus (в клетках крахмал отсутствует) (по Н.В. Первухиной, 1979)
От размера чашелистиков и лепестков, содержания в них хлорофилла зависит ассимиляционная деятельность околоцветника, что влияет на скорость фотосинтеза. Например, по данным Н.В. Первухиной (1979), интенсивность фотосинтеза околоцветника Nymphaea Candida находится в пределах 6,3-9,6 мг СО2 на 1 г сухого вещества за 1 ч, в то время как у листа этого растения данный показатель достиг 42,6- 62,6 мг СО2 на 1 г сухого вещества за 1 ч. У Papaver rhoeos интенсивность фотосинтеза околоцветника практически одинакова с интенсивностью фотосинтеза листьев. Согласно данным Porter et al. (1950), Grundbacher (1963), за счет ассимиляционной деятельности цветковых чешуй создается около 30% урожая. Подобного мнения придерживаются Н. С. Петинов и А. Н. Павлов (1957) которые считают, что к моменту налива зерновки активным остается верхний лист. В это время чешуи являются интенсивно функционирующими органами. Таким образом, ассимиляционная деятельность околоцветника играет существенную роль в формировании семян и плодов.
Кроме зеленых пластид в околоцветнике встречаются хромопласты. Они обусловливают желтую окраску цветков, например у сложноцветных, тыквенных. С. И. Лебедев (1953) обосновал физиологическую роль желтых пластид околоцветника некоторых растений. По его мнению, хромопласты околоцветника ложно-язычковых цветков корзинки подсолнечника участвуют в физиологических процессах, протекающих в генеративной сфере.
Известно, что лепестки у многих растений опадают после оплодотворения цветка. По наблюдениям В. Г. Александрова и др. (1951), клетки опавших лепестков, помещенных во влажную среду, функционируют в течение долгого времени. В опыте Muller (1893) отрезанные от раскрывшегося цветка и помещенные под стеклянный колпак лепестки функционировали дольше, чем лепестки, находившиеся на цветке. Ivanov (1927) считает, что околоцветник обладает высокой интенсивностью транспирации, так как содержит много влаги и имеет тонкостенный эпидермис со слаборазвитой кутикулой. Поэтому растение, предохраняя себя от излишней потери воды, сбрасывает живые лепестки, жертвуя определенным количеством питательных веществ.
После раскрывания цветка существенно увеличивается интенсивность его дыхания. По данным Hsiang (1951), распустившийся цветок дышит в 3 раза интенсивнее, чем бутон. В результате протекающих биохимических процессов в цветке накапливается большое количество токсикантов (эфирные масла, дубильные вещества, кристаллы оксалата кальция), которые сосредотачиваются в околоцветнике. Сбрасывая лепестки, цветок освобождается от конечных продуктов распада, предохраняя себя от самоотравления.
Обладая высокой интенсивностью транспирации, околоцветник играет существенную роль в водном режиме цветка (Ivanov, 1927). Например, в гидроцитную систему эпидермиса цветковых чешуй злаков входят окремневшие бугорки (Л.Р. Петрова, 1958). Наружные стенки этих клеток сильно утолщены, так что образуют бугорок над поверхностью эпидермиса. По краям окремневших бугорков в наружной стенке клетки располагаются крупные поры. Окремневшие клетки формируются над основной гидроцитной тканью чешуи, которая простирается до проводящих пучков (К.Ж. Сыбанбеков, 1965). Таким образом, в теле чешуи образуется единая водопроводящая система. Кроющие клетки эпидермиса цветковых чешуй и остей также пронизаны порами. При этом и наружные стенки клеток имеют большое количество пор. Такой характер строения эпидермальной ткани способствует усилению связи чешуй и остей с внешней средой.
Характерной чертой околоцветника является наличие большого количества волосков на чашелистиках и лепестках. Е.А. Мирославов (1965) при изучении трихом цветка растений из семейства норичниковых выявил особенности этих образований, позволяющие оценить их функциональное значение. Одной из них является раннее заложение и дифференциация трихом околоцветника. Например, зачатки волосков чашелистиков марьянника дубровного возникают в тот период когда еще все клетки, только что заложившегося бутона, носят меристематический характер. Андроцей и гинецей представлены меристематическими бугорками, а цветок не превышает размеры булавочной головки. Деления клеток трихом заканчиваются раньше, чем деления клеток эпидермиса. Дифференциация сосудов начинается после заложения трихом. Еще позднее дифференцируются устьица. Со времени заложения волосков, до полной дифференциации устьичного аппарата, бутон увеличивается в 18-20 раз.
Волоски располагаются по краям пластинки и над жилками. В тех случаях, когда волоски образуют сплошной покров, они распределяются более равномерно по всей поверхности органа. На определенную связь между опушением и проводящей системой указывают Е. И. Проскорякова др. (1929) считают, что при густом опушении сеть жилок развита слабее и, наоборот, при редко расположенных волосках проводящая система становится более мощной.
Высокую концентрацию клеточного сока имеют апикальные клетки трихом. В направлении от верхней части волоска к основанию осмотическое давление в клетках трихом понижается. Увеличение осмотического давления в акропетальном направлении наблюдается и у трихом-гидатод. В литературе также имеются сведения о значительно более высокой кутикулярной транспирации клеток волосков по сравнению с обычными эпидермальными клетками (З. Штруггер, 1953).
Волоски формируются и начинают функционировать как образования, насасывающие и испаряющие воду в период эмбрионального роста органа, когда межклетники развиты слабо, устьица еще не сформированы, цветок закрыт прицветниками. Поэтому деятельность волосков представляет собой существенный фактор, обеспечивающий непрерывный приток воды и питательных веществ к тканям развивающегося цветка. С ростом органа, дифференциацией его тканей, развитием мощной системы межклетников, формированием устьиц, роль волосков снижается. В связи с этим волоски отмирают или, изменяя структуру, выполняют другие функции. Следовательно, трихомы околоцветника, осуществляют важную роль в передвижении воды и солей к тканям цветка (Н.В. Первухина, 1979).
Большое значение для энтомофилии имеет околоцветник. Например, мелкие соцветия некоторых растений (сложноцветные, молочайные) по воздействию на опылителей тождественны более крупным цветкам (эспарцет, гречиха), представляя собой «единицы привлечения». Для наименования такой единицы предложен термин «blossom» (Kugler, 1963).
Функцию привлечения могут осуществлять ярко окрашенные тычинки. Например, соцветия различных видов Mimosa, Acacia привлекают энтомофилов многочисленными окрашенными тычинкам на длинных тычиночных нитях. Связники тычинок могут образовывать крупные и ярко окрашенные придатки (лимонно-желтые, малиновые, красные), заменяющие по функции венчик (М.М. Ильин, 1953).
Большую роль в работе насекомых играет запах цветка. Muller (1893) делает вывод, что запах превосходит значение окраски цветка в привлечении насекомых. Он указывает, что например, пряно пахнущие цветки Convolvulus arvensis посещаются насекомыми охотнее, чем растущие рядом, более заметные, но лишенные запаха цветки С. sepium.
У многих растений между чашелистиками и верхушечными листьями имеются переходы. Например, у Erica vestita, Calluna vulgaris чашелистики по форме и строению сходны с листьями, что может свидетельствовать об их сходной природе (А.Д. Имс, 1964).
По мнению А.А. Федорова и др. (1950) в случаях, когда члены околоцветника отличаются от верхушечных листьев по цвету и форме, как например, у представителей рода Trollius, часто встречаются цветки с отклонениями от нормы.
Согласно нодальной анатомии чашелистики снабжаются проводящей системой подобно листьям, в то время как снабжение лепестков аналогично снабжению тычинок. По мнению Hiepko (1965), независимо от размеров, формы и плотности лепестка, его след составлен одним пучком, как и след тычинки.
По своему заложению и развитию на конусе нарастания чашелистики подобны вегетативным листьям: они обычно закладываются последовательно по спирали, для лепестков же характерно одновременное заложение на конусе нарастания. Почкосложение чашелистиков константно, в противоположность чрезвычайно изменчивому почкосложению лепестков (Goebel, 1933).
Согласно исследованиям А. Л. Тахтаджяна (1964) лепестки развились у большинства покрытосеменных путем стерилизации тычинок. Например, у Magnoliales все части околоцветника морфологически эквивалентны, а если имеется дифференциация на чашечку и венчик, то все части околоцветника брактейной природы (Hiepko, 1965).
Smith (1928) отмечает, что расположение лепестков и тычинок составляет непрерывную спираль. Он считает, что в противоположность чашелистикам, имеющим сходную с листьями проводящую систему, лепестки имеют след, составленный одним пучком (рис. 3). В ряде случаев это различие является следствием того, что в чашелистиках пучок ветвится перед вхождением в орган, а в лепестках - после вхождения в него (рис. 4).
С помощью нодальной анатомии установлено, что оранжево-желтые члены околоцветника Trollius (рис. 5) представлены чашелистиками, стаминодии могут быть отнесены к лепесткам (Hiepko, 1965).
Cutter (1965) отметил тесную физиологическую связь между лепестками и тычинками. Например, рост венчика связан с развитием тычинок, так же как рост чашечки - с развитием плодолистиков. По мнению Heslop-Harrison (1959), ауксин может подавлять рост венчика и тычинок и стимулировать развитие чашечки и гинецея. Было показано, что при удалении тычинок в гермафродитном цветке Glechoma замедлялся рост венчика. При обработке такого цветка гиббереллином венчик восстанавливает свои размеры. Обработка женского цветка гиббереллином ведет к гигантскому увеличению венчика (Plack, 1958).
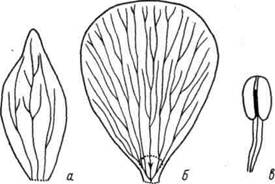
Рис. 3. Жилкование чашелистика (а), лепестка (б) и тычинки (в) Ranunculus repens (по Smith, 1928)
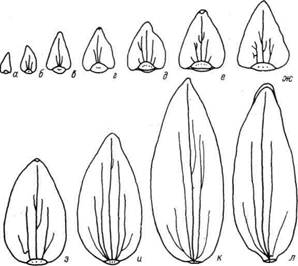
Рис. 4. Элементы околоцветника Nandina domestica. а - предлист; б-и - по одному чашелистику из 1-6-го, 8-го и 10-го кругов соответственно; к, л - лепестки из 12-го и 13-го кругов околоцветника (по Smith, 1928)

Рис. 5. Trollius asiaticus - пример цветка с венчиковидной чашечкой (фото Д.М. Панкова, 2010)
Таким образом, кроме привлечения насекомых-опылителей и защиты цветковых органов от неблагоприятных внешних условий, околоцветник играет важную роль в общем цикле физиологических процессов, протекающих в цветке. Он служит вместилищем запасных питательных веществ, идущих на построение плодов и семян, принимает участие в фотосинтетических процессах, является местом накопления продуктов метаболизма. Летучие выделения околоцветника способствуют прорастанию пыльцы. Существенное значение околоцветник имеет в водном режиме цветка.