
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

5.3.4.Общая характеристика развития органов пищеварения
По происхождению пищеварительная система является наиболее древней и массивной по объему, площади переваривания и всасывания, служит для захвата пищи, определения ее вкусовых качеств, пережевывания, проглатывания, химического переваривания, всасывания, питательных веществ и воды в кровь, лимфу и выделения непереваренных пищевых масс наружу. Она пронизывает тело коз спереди назад, по этой причине, располагается во всех полостях тела.
У коз, как и у всех млекопитающих, условия внутриутробного развития обеспечиваются формированием, так называемых провизорных (временных) органов, в состав которых входят желточный мешок, амнион, аллантоис, хорион и плацента. В образовании плаценты принимают участие провизорные органы и слизистая оболочка матки. Из данного анализа следует, что питательные вещества в зародыш поступают путем всасывания через трофобласт «маточного молочка» желез слизистой оболочки матки – это единственный источник питания в этот период его развития.
Предплод и ранний плод питательные вещества получают через плаценту из крови матери и через стенки водных оболочек из околоплодной жидкости. Позднего плода, к двум последним источникам добавляется пищеварительная система, которая со второй половины плодного периода развития начинает заглатывать и переваривать амниотическую жидкость.
Основываясь на смене внутриутробного питания и развития зародыша, предплода и плода ученые разработали периоды их развития, в том числе и критические.
Зародышевый период коз продолжается 1–29 дней и характеризуется дроблением, образованием зародышевых листков, формированием зачатков провизорных органов (трофобласта, желточного мешка, амниона, аллантоиса), ранним органогенезом, в том числе и пищеварительной системы, которая закладывается из внутреннего зародышевого листка.
Предплодный период (30–45 дней) С.Н. Боголюбский (1968) относит к переходному развитию от зародыша к плоду. Этот период характеризуется развитием плаценты и началом формирования органов, систем организма. С формированием плаценты заканчивается лактотропное развитие зародыша и наступает гематотропное питание предплода, плода и продолжается до рождения.
В течение всего плодного периода развития плацента постоянно усложняется и достигает максимального развития к рождению, происходит дифференциация органов и их функции, меняются отношения между матерью и плодом, что послужило причиной разделения его на ранне – (1,5–3,5 мес.) и позднеплодный периоды, продолжающийся до родов.
В целом их пищеварительная система подразделяется на головной отдел или ротоглотку, пищевод и желудок, тонкий (средний), толстый (задний) отделы. Следует отметить, что она у коз исследована недостаточно, особенно, формирование, развитие, как органов, так и их гистоструктур в онтогенезе. Нет работы по иннервации и кровоснабжению, изменению функции с возрастом, что имеет большое практическое и теоретическое значение.
Развитие ротоглотки
Ротоглотка пищеварительной системы подразделяется на рот и глотку. В целом органы головной кишки изучены в самых общих чертах, но более полно описана смена зубов коз.
В трехлистковую стадию развития зародыша, на его головном конце, снизу мозговых пузырей, за счет впячивания наружного зародышевого листка назад появляется ротовая ямка, а навстречу ей из внутреннего зародышевого листка формируется кишечная бухта (рис. 22). До возраста козлят 33–35 дней их разделяет глоточная перепонка (первичное небо). В дальнейшем перепонка исчезает (прорывается). Из ротовой ямки формируются рот и его органы, а из кишечной бухты – глотка, пищевод.
Поэтому эпителиальная выстилка рта, ротоглотки и гортанной части глотки, железы, эмаль зубов – эктодермального происхождения. Однако не ясно, каким образом многослойный плоский эпителий стал выстилать пищевод и даже преджелудки коз. Это вопрос ждет своего разрешения.
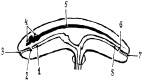
Рис. 22. Схема продольного разреза раннего предплода:
1 – кишечная бухта; 2 – глоточная перепонка; 3 – ротовая ямка; 4 – мозговые пузыри; 5 – спинной мозг; 6 – анальная перепонка; 7 – анальная ямка;
8 – анальная бухта
Развитие языка происходит сложно. На ранней стадии предплода, в области образования рудиментов жаберных дуг и карманов, возникают непарный бугорок и два парных валика (рис. 23).
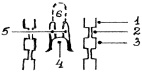
Рис. 23. Схема развития языка:
1 – первая жаберная дуга; 2 – второй жаберный карман; 3 – вторая жаберная дуга; 4 – непарный бугорок; 5 – парные валики; 6 – тело и верхушка языка
Непарный бугорок закладывается по одним данным в области вентральных отделов I и II (А.И. Акаевский, 1975), по другим – II, III и IV жаберных дуг (В.Н. Жеденов, 1965). Парные валики эктодермального происхождения охватывают справа и слева непарный бугорок, несколько выдвигаясь вперед, образуют впоследствии тело и верхушку языка, а непарный бугорок превращается в корень языка.
Застенные железы рта
К застенным железам ротоглотки относят околоушную, челюстную и подъязычную железы. Формируются они путем впячивания эпителия ротовой ямки в ее стенку, по типу образования экскреторных желез. Раньше возникает околоушная железа, затем челюстная и подъязычная. Окончательно железы заканчивают формирование после рождения (В.Н. Жеденов, 1941).
Околоушная железа
Железа бледнорозового оттенка, представляет компактный, трубчато-альвеолярный, дольчатый орган, расположенный между задним краем большой жевательной мышцы и ярёмным отростком. Её верхний край окаймляет основание наружного слухового прохода, а нижний – достигает угла ветви челюсти. Внутренней поверхностью железа прикрывает большую жевательную мышцу, височно-челюстной сустав, околоушной лимфатический узел и верхнюю челюстную вену. Снаружи железа покрыта поверхностной фасцией, околоушнораковинной мышцей, подкожной мышцей головы и частично – щёк.
Из нижнего переднего угла железы выходит околоушной проток, идёт вперед дугообразно по нижней трети края большой жевательной мышцы, сопровождая лицевую артерию и вену. Проток поднимается до отхождения глубокой лицевой артерии и вены. Здесь он поворачивает вперёд и на уровне III–IV коренного зуба взрослых коз открывается на сосочке в защёчное пространство ротовой полости.
С возрастом коз проток в защёчное пространство открывается не одинаково: так у новорожденных и 20-дневных козлят – на уровне середины II молочного премоляра; в месячном и трёхмесячном возрасте – на уровне II–III молочного премоляра; IV–VI-месячном возрасте – на уровне III молочного премоляра, а у половозрастных – на уровне III премоляра и I моляра.
Изменение места открытия с возрастом протока в защёчное пространство ротовой полости обусловлено, на наш взгляд, ростом, развитием и сменой молочных зубов на постоянные, появлением постоянных моляров и развитием жевательных мышц.
Диаметр протока новорожденных козлят равен 0,12 мм, трёх и четырёх- месячных – 0, 51 мм, годовалых и трёхлетних – 2,88 мм и 4-летних – 3,14 мм.
Масса околоушных желез с возрастом плодов и коз после рождения возрастает неравномерно, причём левой железы незначительно превалирует над правой. Так, у трёхмесячных плодов-козочек масса левой железы составляет 0,8 и правой – 0,7 г, а у козликов – 0,9 и 0,8 г. У четырёхмесячных плодов масса желез уже достигает, соответственно 1,3 и 1,1; 2,2 и 1,9 г. На момент рождения масса желез несколько понижается и у новорожденных равняется: у козочек – 0,5 и 0,4 г, а у козликов – 0,8 и 0,7 г. Спустя 20 дней после рождения отмечается увеличение массы желез как у козочек, так и у козликов, соответственно в 2,18 и 1, 9; 3,4 и 3,7 раза. Волнообразно масса желез возрастает до 36-месячного возраста: у козочек достигает правая 16,8 и левая 17,6 г, козликов 18,7 и 18, 9 г. В пятилетнем возрасте достоверно наступает снижение массы желез, что свидетельствует о наступлении инволютивных изменений.
Нижняя челюстная железа
Железа желтоватого оттенка, имеет чёткое дольчатое строение и более компактна, чем околоушная, иеет вытянутую грушевидную форму с округлыми краями, размещается в межчелюстном пространстве. Внутренней поверхностью прилежит к глотке и гортани, а наружной – к медиальной крыловидной мышце и к ветви нижней челюсти. Сзади и вверх железа достигает переднего края крыла атлента, снаружи прикрыта латеральным заглоточным лимфатическим узлом.
Проток выходит из переднего угла железы на уровне сосудистой вырезки нижней челюсти, тянется вперёд по наружной поверхности боковой язычной мышцы, затем по нижнему краю однопротоковой подъязычной железы и открывается на подъязычной бородавке, под верхушкой языка.
Диаметр протока у новорожденных равен 0,08 мм, шестимесячных – 0,23 мм, 12–36-месячных – 1,17 мм и у пятилетних – 1,54 мм.
Масса левой и правой железы плодов-козочек в три месяца достигает 0,9 и 0,7 г, в четыре месяца увеличивается до 1,6 и 1,4 г, соответственно. После рождения масса до 20 – дневного возраста уменьшается и в месячном возрасте равна 2,55 и 2,25 г. В три месяца левая железа достигает 4,5, а правая – 4,2 г. Наиболее интенсивно массы железы возрастает с четырёх до шестимесячного возраста. Практически она утраивается, и достигает, соответственно 14,8 и 14,3 г. В три года она равна уже 21,8 и 19,6 г. Из данного анализа следует, что масса нижней челюстной железы значительно больше, чем околоушной и увеличивается неравномерно. При этом замечено, что масса железы козликов выше, чем у козочек и левая железа преобладает по массе над правой железой.
Подъязычная железа
Подъязычная железа коз желтоватовосковидного цвета делится на однопротоковую и многопротоковую. Обе части располагаются в боковом подъязычном кармане ротовой полости, в подъязычной складке слизистой оболочки дна рта.
Однопротоковая компактнее многопротоковой железы и имеет ланцетовидную форму. Она лежит несколько ниже и наружу от многопротоковой железы и простирается от подбородка до предпоследнего коренного зуба нижней челюсти. Выводной проток её идёт параллельно с протоком нижней челюстной железы и открывается на подъязычной бородавке.
Многопротоковая железа лежит поверхностнее длиннопротоковой и простирается от середины межальвеолярного края нижней челюсти до корня языка. Она состоит из большого числа железистых долек соединенных между собой рыхлой соединительной тканью. Её многочисленные выводные протоки открываются в боковой подъязычный карман дна ротовой полости.
Масса подъязычной железы в плодном периоде развития возрастает неравномерно и к рождению левой достигает 0,67 и правой 0,51 г. В возрасте козлят одного месяца после рождения, соответственно, равняется 0,83 и 0,61 г. Максимума масса железы достигает у коз в возрасте 36 месяцев и составляет 7,7 и 7,12 г. К 60-месячному возрасту масса железы уменьшается, соответственно на 2,5 и 2,0 г. У козликов во всех возрастных периодах масса железы была выше, чем у козочек.
Из анализа развития застенных желез ротоглотки следует, что масса всех желез увеличивается неравномерно как у козочек, так и у козликов. Наиболее интенсивно она увеличивается с 3 месяцев постнатального периода развития и до 12 месяцев, что на наш взгляд совпадает с началом перехода от молочного питания к питанию грубыми кормами, свойственному взрослым козам. В плодном и постнатальном периоде развития масса нижней челюстной железы возрастает интенсивнее, чем околоушной и подъязычной желез. Наряду с этим масса всех застенных желез козликов на всех этапах развития больше, чем козочек и масса всех левых – как козликов, так и козочек больше, чем правых.
Биологические активные вещества застенных желез рта
К биологическим активным веществам застенных желез рта относят: калликреин, гистамин, ренин, тонин, паротин, кальцитониноподобное вещество, фактор роста нервов (ФРН), эпидермиса (ФРЭ), мезодермы (ФРМ), летальности (ФЛ) и др. В настоящее время изучена молекулярная биология ФРН. ФРЭ, но ряд биологически активных веществ желез остаются не изученными. Эти вещества не только влияют на кровоснабжение, трофику, секрецию и функцию органов, но поступая в кровеносное русло, могут повышать или понижать давление крови в организме.
Калликренин обладает гипотензивным фактором, его много в слюне нижней челюстной железы и мало в околоушной и подъязычной железе. Это протеолитический фермент обладающий свойствами сериновых протеиназ.
Ренин и тонин выделен из слюны и ткани нижней челюстной железы.
Ренин – фермент, который обладает сосудосуживающим действием и стимулирует выделение альдостерона. Последний синтезируется в основном в коре надпочечников, регулирует водный, минеральный обмен в организме, главным образом катионов Na, K.
В настоящее время установлено, что наряду с почками, ренин синтезируют застенные железы рта, семенники, клетки головного мозга и аденогипофиз (О.И. Сукманский, 1991). Учёные предполагают, что внепочечный ренин регулирует кровоток и воспаления в очагах поражения, предотвращает проницаемость стенок сосудов, заживляет раны и т.д.
Продуктом ренина является ангиотензин, участвует в регуляции артериального давления, водно-солевого баланса и влияет на секреторную функцию застенных желез рта.
Установлено, что ренин синтезируется в зернистых протоках нижней челюстной железы, а концентрируется в секреторных гранулах вместе с ФРЭ. В связи с этим, О.И. Сукманский указывает, что высокая активность ренина обнаруживается в очищенных препаратах фактора роста нервов.
Тонин – фермент, который содержится в большом количестве в нижней челюстной железе. По действию тонин сходен с ренином и способен переходить в ангиотензин. Тонин из железы выделяется легко в слюну и в оттекающую венозную кровь. В настоящее время выдилен в очищенной форме. Сходство его действия с ренином, объясняется отношением тонина к числу сериновых протеиназ. Но физиологическая роль его до конца ещё не выяснена, способствуя образованию ангиотензина, может влиять на артериальное давление, водно-солевой баланс организма.
Фактор роста нервов (ФРН) выделяется в слюне нижней челюстной железы и обнаружен в её ткане, стимулирует деление, дифференцировку и рост аксонов симпатических нейронов ганглиев.
Гистамин – это гетероциклическая аминокислота гистидина. Содержится в тканях застенных желез рта и слюне, высвобождается в больших количествах во время аллергических реакций, расширяет кровеносные сосуды, сокращает гладкую мышечную ткань, повышает секрецию свободной хлористоводородной кислоты железами желудка.
Фактор роста эпидермиса (ФРЭ) – полипептид, выделяемый нижней челюстной железой, способствует преждевременному открытию глаз, прорезыванию зубов у новорожденных и тормозит секрецию желез желудка. При введении животным способствует уеличению толщины эпидермиса, истончению основы кожи и подкожного слоя.
Фактор роста мезодермы (ФРМ) был открыт сравнительно недавно (О.И. Сукманский, 1991). Представляет гомогенат слюнных желез, состоящий из двух фракций: первая – это тимоцитотрансформирующий фактор, а вторая – две фракции роста клеток соединительной ткани, которые были названы фактором роста мезодермы (V.L. Weimar, E.L. Sguires, R.J. Knoch, 1975). ФРМ стимулирует развитие и деление фибробластов в тканевой культуре, активирует деление и миграцию эндотелиальных клеток при повреждении и ускоряет заживление ран роговицы глаза.
Паротин является активным началом околоушной железы. В сухом виде может долго сохраняться, а в растворе быстро инактивируется, в том числе в желудочно-кишечном тракте. Поэтому его вводят в организм внутримышечно. В Японии производят в промышленных масштабах. Применяют при лечении пародонтоза, ксеростомии (xeros – сухой, stomatos – рот), при дистрофических поражениях костей, суставов. Паротин и его аналоги, введённые животным, способствуют развитию, росту костей, эмали и дентина зубов, и их обызвествлению, увеличивает количество лейкоцитов в крови. Повышает интенсивность обмена фосфора, натрия в зубах.
Смена зубов
Зубы коз относятся к смешанному типу: резцовые – короткокоронковые, премоляры, и моляры – длиннокоронковые, лунчатые. Резцовые и премоляры имеют молочных предшественников, моляры нет.
Резцовые зубы имеются только на нижней челюсти. Они подразделяются на зацепы, расположенные справа и слева от стреловидной (сагиттальной) плоскости, затем внутренние, наружные – средние и окрайки. Всего их четыре, клыков – нет, молочных и постоянных премоляров на каждой стороне челюсти коз – три, моляров тоже три. Из этого следует, что у молодых коз молочных зубов всего 20, а постоянных – 32 зуба.
Определение возраста коз разработано по смене и стиранию резцовых зубов. Козлята на белый свет появляются с молочными зацепами и внутренними средними зубами, на 18–23 день после рождения вырастают наружные средние и окрайки. Таким образом, на 24 день после рождения у развитых козлят имеются все молочные резцовые зубы.
Молочные зацепы меняются на постоянные в возрасте коз
14–16 месяцев, внутренние средние – в 20–22, наружные средние – в 24 и окрайки – в 32–34 месяца. Из этого следует, что к трехлетнему возрасту коз молочные резцовые зубы меняются на постоянные. Ширина жевательной поверхности постоянных резцовых зубов молодых коз достигает 8 мм.
На третьем году жевательная поверхность зубов остается узкой, в четыре года на фоне ее белой поверхности появляется желто-коричневая полоска, и корни зубов выдвигаются из альвеол в виде узкой линии, шириной до 2 мм. В пять лет, в связи со стиранием зубов, желто-коричневые полоски светлеют (А.Д. Шевченко, 2007)
В 6–7 лет корни зубов выдвигаются из альвеол на 3–4 мм, и жевательная поверхность косо вытягивается назад, на всю их язычную поверхность, приобретая четырехгранную или округловытянутую форму. На этой поверхности появляются черные луночки и желтые полоски. В 7–7,5 лет корни зубов над деснами возвышаются не менее чем на 5 мм. Однако, работ по смене предкорневых и стиранию коренных зубов коз в литературе мы не встретили.
Замечено, что у новорожденных козлят уже имеются молочные премоляры. В возрасте 17–22 дней они начинают жевать и поедать комбикорм, отдельные стебли зеленых побегов, а в месячном возрасте они уже развиты, что способствует питанию грубыми кормами.
Развитие глотки
Г.А. Гиммельрейх (1958–1981) делит глотку на носоглотку, ротоглотку и гортанную часть глотки. Пищеварительный путь проходит через ротоглотку, гортанную часть в пищевод, а дыхательный – через носоглотку, гортанную часть глотки в щель гортани и далее в дыхательное горло. Из этого следует, что гортанная часть глотки является общей для дыхательного и пищеварительного путей.
В глотку открываются 7 отверстий: зев, два отверстия хоан, два отверстия слуховых труб, переднее пищеводное отверстие и гортанная щель, ведущая через гортань в дыхательное горло (трахею).
Стенка глотки представлена слизистой мышечной и наружной соединительнотканной оболочками. Слизистая оболочка носоглотки покрыта многорядным мерцательным эпителием, ротоглотки и гортанной части и глотки – многослойным плоским эпителием. Мышечная оболочка дифференцирована на передний, средний, задний констрикторы (суживатели) и на расширитель глотки. В стенке ротоглотки заложена небная миндалина, а справа и слева от надгортанника – околонадгортанная миндалина.
Глотка образуется из жаберной области кишечной бухты. Передний конец кишечной бухты оканчивается у ранних предплодов глоточной перепонкой, после ее прорыва, она соединяется с ротовой ямкой. В итоге появляется сообщение. По этой причине считается, что эпителий рта, ротоглотки и гортанной части глотки эктодермального происхождения.
В стенках правой и левой стороны, будущей глотки, появляются жаберные карманы, а между ними четыре дуги (рис. 24). Первая жаберная дуга превращается в челюстную, вторая – в подъязычную дугу, III и IV – в подъязычную кость и щитовидный хрящ
гортани.
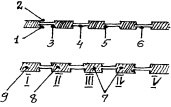
Рис. 24. Схема развития глотки:
I–V – жаберные дуги, а между ними находятся карманы:
1 – закладка слуховых труб и полости среднего уха; 2 – закладка наружного уха; 3 – образование щитовидной железы; 4 – зачаток глоточных миндалин; 5 – закладка паращитовидных желёз и 6 – вилочковой железы;
7 – из IV–VI жаберных дуг происходит щитовидный хрящ;
8 – из 11-З дуг – подъязычная кость;
9 – I дуга превратится в нижнюю челюсть.
Из первого жаберного кармана (брызгальце акул), с его внутренней борозды, развиваются слуховые трубы и полость среднего уха, из внешней – наружный слуховой проход, а разделяющая их мембрана, превращается в барабанную перепонку. Из второго кармана закладываются миндалины и щитовидная железа, из III и IV – тимус и околощитовидная железа.
Развитие пищевода
Пищевод – перепончатомышечная трубка, масса которого у новорожденного козленка составляет 4,6 – 6,7 г, в месячном возрасте – 8,5 г, в годичном – 29,0 г и в возрасте трех лет – 50,0 г. Наиболее интенсивный абсолютный прирост массы пищевода отмечается в возрасте 12-месяцев коз, затем темп прироста несколько падает, но продолжает постепенно увеличиваться до трех лет.
Длина пищевода медленнее возрастает, чем его масса (табл. 69). Так, она в три года достигает 59,0 см и увеличивается по сравнению с новорожденными почти в три раза. Абсолютный прирост длины пищевода увеличивается до месячного возраста, затем темп прироста несколько уменьшается, но высоким сохраняется до одного года. Старше года прирост длины постепенно снижается до трех лет.
Таблица 69
Динамика роста массы и длины пищевода
|
Промеры |
К-во исследований |
Возраст |
|||||
|
Новорожденные |
1 |
6 |
12 |
36 |
72 |
||
|
Масса, г |
4 |
6,3 |
8,8 |
29,0 |
32,0 |
50,0 |
54,0 |
|
Прирост |
4 |
– |
2,5 |
20,2 |
3,0 |
18,0 |
4,0 |
|
Длина, см |
4 |
16,0 |
31,0 |
37,4 |
58,1 |
59,0 |
51,0 |
|
Прирост |
4 |
– |
15,0 |
6,4 |
20,7 |
0,9 |
2,0 |
На всем протяжении стенка пищевода коз тонкая, в шейной части образуется небольшое ампулообразное расширение, которое в области первых ребер сужается, в грудной части вновь расширяется, а в пищеварительном отверстии диафрагмы и преддверии рубца несколько зауживается. Из данного анализа следует, что пищевод на протяжении от глотки до рубца образует два ампулообразных расширения, а между ними одно сужение. Об этом необходимо помнить специалистам при скармливании козам корнеклубнеплодов, так как может быть закупорка пищевода.
Развитие желудка
Желудок у мелкого рогатого скота закладывается из первичной кишки на 20 день зародышевого развития (О.Б. Шумкина, 1960). В начале на короткий срок формируется однокамерный желудок, к 22 дню – он становится двухкамерным, к 25 – трехкамерным и в 27 дней – четырехкамерным. Из анализа следует, что исходной формой для мелких жвачных является однокамерный желудок (Л.В. Давлетова, 1974), который, затем, трансформируется в многокамерный, состоящий из рубца, сетки, книжки и сычуга.
В течение раннеплодного периода развития происходит неравномерность роста органов пищеварения. В этот период интенсивно развивается печень, а желудок и кишечник в росте объема, массы отстают, но к концу периода печень в росте уступает быстро развивающемуся желудку и кишечнику. В течение всего плодного периода развития интенсивность роста рубца, уступает сычугу, что, особенно, заметно к концу плодного периода. Поэтому у новорожденных козлят сычуг относится к рубцу – как 2:1.
В начале плодного периода развития наибольшим морфофункциональным изменениям подвергается слизистая оболочка сычуга. В ней происходит закладка гастронов, из которых в будущем формируются пищеварительные железы. Гастрон – это крипта и впадающий в нее проток, формирующейся железы. К концу раннеплодного периода, таким образом, образуется кардиальная, донная и пилорическая зоны желез. По мере дифференциации гастронов в эпителиальных клетках уменьшается количество гликогена, но увеличивается нейтральных мукополисахаридов, что указывает на активное участие желудка ранних плодов в углеводном и жировом обмене.
В рубец открывается пищевод. Место открытия пищевода в рубец носит название преддверия рубца, внутри его, на границе с сеткой, располагается пищеводный желоб. Во время приема жидкой пищи губы пищеводного желоба смыкаются в трубку и пища из пищевода, минуя преджелудки, проходит в сычуг. Нормальный глоток молока у козленка – сосуна равен 1,5–3,0 мл. В случае продолжительного голодания и жажды козлята с жадностью сосут матерей и глоток, при этом, увеличивается в объеме на два и более раза. В этом случае губы пищеводного желоба не выдерживают напряжения глотка, размыкаются и молоко выплескивается в рубец. Под действием микрофлоры рубца оно сбраживается, загнивает и продукты распада вызывают интоксикацию организма и гастроэнтерит, что приводит к падежу. Поэтому, надо следить за режимом кормления козлят-молочников.
Масса желудка в 12 месяцев достигает 1925,0 г, в три года – 3379,0 г, к 7 годам она увеличивается незначительно. Масса рубца была во всех возрастных группах выше, чем сычуга, а сетки – чем книжки.
Абсолютный прирост массы желудка интенсивно возрастает до трех лет, затем темп прироста снижается. Однако высокий темп наблюдается до года и достигает 1340 г, а в последующие два года 1450 г. До года по приросту массы выходит на первое место рубец, затем сычуг, сетка и книжка. В последующие возрастные периоды происходит увеличение их массы и прироста, но значительно и ниже, чем до года (табл. 70).
В относительных величинах масса желудка наиболее резко увеличивается у козлят до возраста шести месяцев, затем темп роста до трех лет снижается.
У мелких жвачных в новорожденном периоде меньшего развития достигает желудок (Л.В. Давлетова, 1974), особенно преджелудки, а из них – рубец. Об этом свидетельствуют полученные данные (табл. 71).
Объем преджелудков коз с возрастом увеличивается неодинаково, у новорожденных объем сычуга превосходит рубца в 2,3 раза, а книжка сетку – в 1,1 раза, в месячном возрасте козлят рубец уже больше по объему сычуга – в 3,2 раза, а сетка превосходит книжку – в 2,8 раза. С месячного возраста козлят и старше по объему роста на первое место выходит рубец, затем располагаются последовательно сычуг, сетка и книжка. Рост объема преджелудков, как и всего желудка, устанавливается в три года и достигает в среднем, 17238,0 мл.
Абсолютный прирост объема преджелудков и всего желудка коз наиболее интенсивно увеличивается в 6 месяцев, затем постепенно понижается до трех лет. Однако темп абсолютного прироста объема желудка по возрастам неодинаков, так до месячного возраста он увеличивается в 8,5 раз, в 6 месяцев – 5,9, а в 36 – в 1,8 раза ё (табл. 71).
Из данного анализа следует, что морфологически многокамерный желудок коз функционально готов переваривать грубую пищу в возрасте козлят одного месяца, хотя грубый корм они начинают поедать с 18–22 дней.
Таблица 70
Динамика возрастных изменений массы желудка
|
Возраст, мес. |
Масса, г |
Абсолютный прирост, г |
Относительный рост, раз |
||||||||||||
|
Рубец |
Сетка |
Книжка |
Сычуг |
Всего |
Рубец |
Сетка |
Книжка |
Сычуг |
Всего |
Рубец |
Сетка |
Книжка |
Сычуг |
Всего |
|
|
Новорожденные |
38,0 |
8,2 |
2,9 |
21,0 |
70,1 |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
|
1 мес. |
48,0 |
11,2 |
3,9 |
27,5 |
90,6 |
10,0 |
3,0 |
1,0 |
6,5 |
20,5 |
1,26 |
1,36 |
1,34 |
1,31 |
1,3 |
|
6 мес. |
375,0 |
64,2 |
62,5 |
64,0 |
577,7 |
327,0 |
53,0 |
58,6 |
36,5 |
487,1 |
7,8 |
5,7 |
16,0 |
7,3 |
6,4 |
|
12 мес. |
1342,5 |
72,5 |
70,0 |
433,0 |
1925,0 |
967,5 |
8,3 |
7,5 |
369,0 |
1347,3 |
3,6 |
1,1 |
1,1 |
6,8 |
3,3 |
|
36 мес. |
2050,0 |
98,0 |
81,0 |
1150,0 |
3379,0 |
707,5 |
23,5 |
11,0 |
717,0 |
1454,0 |
1,52 |
1,4 |
1,2 |
2,6 |
1,8 |
|
72 мес. |
3535,0 |
122,5 |
100,0 |
1314,0 |
3971,5 |
485,0 |
24,5 |
19,0 |
64,0 |
592, |
1,23 |
1,3 |
1,2 |
1,0 |
1,2 |
Таблица 71
Динамика возрастных изменений объема желудка
|
Возраст, мес. |
К-во |
Объем, мл |
Абсолютный прирост, мл |
Относительный рост, разах |
||||||||||||
|
Рубца |
Сетки |
Книжка |
Сычуга |
Всего |
Рубца |
Сетки |
Книжка |
Сычуга |
Всего |
Рубца |
Сетки |
Книжка |
Сычуга |
Всего |
||
|
Новорожденные |
5 |
35,0 |
8,3 |
9,3 |
80,0 |
132,6 |
? |
? |
? |
? |
? |
? |
? |
? |
? |
? |
|
1 мес. |
3 |
821,0 |
36,6 |
13,0 |
258,3 |
1129,0 |
786,0 |
28,3 |
3,7 |
178,0 |
996,4 |
23,4 |
6,5 |
1,4 |
3,2 |
8,5 |
|
6 мес. |
4 |
5700,0 |
236,0 |
55,0 |
653,0 |
6644,0 |
4879,0 |
199,4 |
42,0 |
394,7 |
5515,0 |
0,69 |
1,6 |
4,2 |
2,5 |
5,9 |
|
12 мес. |
3 |
8250,0 |
371,0 |
120,0 |
740,0 |
9481,0 |
2550,0 |
135,0 |
65,0 |
87,0 |
2837,0 |
1,5 |
2,6 |
2,2 |
1,1 |
1,4 |
|
36 мес. |
3 |
14495,0 |
980,0 |
208,0 |
1555,0 |
17238,0 |
6245,0 |
609,0 |
88,0 |
815,0 |
7757,0 |
1,8 |
? |
1,7 |
2,1 |
1,8 |
|
72 мес. |
3 |
13520,0 |
950,0 |
325,0 |
1558,0 |
16353,0 |
? |
? |
117,0 |
3,0 |
? |
? |
? |
1,6 |
1,0 |
? |
|
По В.Н. Жеденову (1965) |
15-23 |
1-2 |
0,9 |
3,3 |
? |
? |
? |
? |
? |
? |
? |
? |
? |
? |
? |
|
Гистология стенки рубца
Стенка рубца 60-дневных плодов состоит из слизистой, мышечной и серозной оболочек.
Эпителиальное поле слизистой оболочки представлено двумя слоями клеток. Самый глубокий базальный слой состоит из двух-четырёх рядов клеток с интенсивно окрашенной цитоплазмой, в которой располагаются мелкие ядра. Поверхностный слой представлен крупными (пузырчатыми) клетками с большими ядрами и слабо окрашенной цитоплазмой. Слои клеток чётко отграничены друг от друга.
Соединительнотканная основа слизистой оболочки по толщине уступает эпителиальной выстилки. В ней встречаются слабо дифференцированные клетки и другие элементы.
Мышечная оболочка тонкая и имеет слой миобластических элементов, отделяющихся друг от друга прослойкой. В будущем из неё сформируется циркулярный и продольный слои. Серозная оболочка относительно толстая и содержит малодифференцированные мезенхимные элементы, и разреженные сосудистые сети.
Слизистая оболочка стенки рубца 75-дневных плдодов несколько утолщается, в мышечной оболочке продолжается дифференциация миобластических элементов, значительно утолщается циркулярный слой, особенно в местах формирования мышечных тканей, между слоями появляются кровеносные сосуды, но серозная оболочка, по сравнению с 60-дневными плодами, становится тоньше.
Поверхность эпителиального поля слизистой оболочки 90-дневных плодов остаётся ровной, но ростковый слой уже представлен двумя рядами клеток. Клетки пузырчатого слоя уменьшаются в размерах, изменяется их форма, клетки, прилегающие к ростковлому слою, округляются и к периферии утолщаются. Соединительнотканная основа становится в три-четыре раза тоньше эпитьелиального слоя, в отдельных участках она вдаётся в эпителий, образуя сосочки. Утолщается и уплотняется мышечная оболочка, становится тоньше соединительноьакнная прослойка, разделяющая циркулярный и продольный слои. Серозная оболочка тонкая и покрыта уже мезотелием.
В возрасте плодов 105 дней поверхностные эпителиобласты становятся плоскими, а глубже лежащие уменьшаются в размерах. Соединительнотканные сосочки увеличиваются в высоту, мышечная оболочка продолжается утолщаться, а серозная – истончается.
В возрасте плодов 120 дней на поверхности слизистой оболочки между сосочками появляются пузырчатые клетки, дно выемок становятся плоскими, а прилежащие к сосочкам клетки остаются объемными. Ростковые клетки лежат в ряд, но в местах расположения сосочков они становятся двухслойными. В соединительнотканной основе слизистой оболочки увеличивается число эластических и коллагеновых волокон. Мышечная оболочка четко подразделяется на циркулярный и продольный слои, лишь без видимых изменений сохраняется серозная оболочка.
У 135 дневных плодов на поверхности эпителиального поля слизистой оболочки рубца появляется слой отмирающих клеток, ростковый слой представлен одним рядом призматических клеток, увеличивается общая толщина мышечной оболочки.
Стенка рубца новорожденного козленка представлена уже развитой слизистой, мышечной и серозной оболочками. Пузырчатые клетки эпителиального поля по объему становятся меньше и их ядра занимают почти две трети площади цитоплазмы, со стороны полости рубца слизистая оболочка покрыты слоем мертвых клеток. Базальный слой – везде представлен одним рядом призматических клеток, хорошо развиты сосочки. Заметно утолщается циркулярный слой мышечной оболочки.
У козлят в возрасте одного года сосочки рубца развиты и свободно выдаются в его полость, снаружи покрыты слоем плоских клеток с мелкими ядрами, над ними лежат пузырчатые клетки, с хорошо развитой цитоплазмой и крупными ядрами, а ростковые клетки расположены в один ряд. По размерам они уступают плоским и шиповатыми клеткам. Соединительнотканная основа слизистой оболочки в три раза толще эпителиального слоя. Мышечная оболочка развита, циркулярный слой в 1,5 раза толще продольного. Между слоями развиты сосудистые сети, их ячеи вытянуты по ходу мышечных волокон. Несколько утолщена серозная оболочка, в ее основе мало клеточных элементов.
В двухлетнем возрасте хорошо развиты сосочки рубца, соединительнотканная основа слизистой оболочки становится толще эпителиального поля. Циркулярный и продольный слои мышечной оболочки становятся одинаковыми по толщине. Висцеральная брюшина остается без изменения.
В последующих возрастных периодах изменения в сетке рубца незначительные. Из этого следует, что двухлетнему возрасту коз гистогенез в стенке рубца заканчивается.
Гистогенез стенки сетки
Стенка сетки 60-дневных плодов подразделяется на слизистую, мышечную и серозную оболочки. Слизистая оболочка, в свою очередь, подразделяется на эпителиальный слой и соединительнотканную основу.
Эпителиальное поле также делится на недифференцированный базальный и слой пузырчатых клеток. Ростковый слой клеток лежит на базальной мембране и представлен 2–4 рядами мелких призматических клеток с интенсивно окрашенной цитоплазмой. Ядра их крупные, хорошо выражены. Пузырчатый слой состоит из клеток с цитоплазмой и с крупными ядрами. Форма клеток чаще округлая.
Соединительнотканная основа слизистой оболочки по толщине соответствует эпителиальному слою, в ней неравномерно располагаются слабо дифференцированные клеточные элементы, их больше около базальной мембраны. В основе уже развита сосудистая сеть.
Мышечная оболочка тоньше соединительнотканной основы и четко подразделяется на внутренний циркулярный и наружный продольный слои. Циркулярный слой в 3 раза толще продольного. Между слоями располагается сосудистая сеть. Серозная оболочка тонкая и представлена малодифференцированными клетками.
У 75-дневных плодов слой пузырчатых клеток эпителия слизистой оболочки в четыре-пять раз толще росткового.
Соединительнотканная основа оболочки образует в сторону эпителия выпячивания. В местах выпячиваний, у базальной мембраны, увеличивается количество клеток фибробластического ряда. По толщине она становится равной циркулярному мышечному слою. В этом возрасте циркулярный слой почти в два раза толще продольного мышечного слоя. Серозная оболочка существенно не изменяется.
Эпителиальный слой слизистой оболочки 90-дневных плодов утолщается за счет увеличения толщины пузырчатого слоя клеток, последний по толщине превосходит все слои оболочки в 3,5 раза. Со стороны полости сетки пузырчатый слой клеток начинает уплощаться, на поверхности эпителия появляются лоскуты мертвых клеток, особенно, их много в лакунах между выпячиваниями. Базальный слой клеток уже представлен одним слоем призматических клеток.
Соединительнотканные выпячивания основы слизистой оболочки уже образуют первичные складки, появляется выраженная
четкость, в связи с чем, основа оболочки становится в 10 слишним раз тоньше эпителиального поля.
Мышечная оболочка стенки сетки в целом утолщается, продольный слой остается толще, чем циркулярный, миобластические элементы уплотняются, между слоями хорошо уже выражена сосудистая сеть. Основа серозной оболочки покрыта мезотелием.
Поверхностные пузырчатые клетки эпителия слизистой оболочки 105-дневных плодов становятся плоскими, и на отдельных его участках происходит отторжение их в просвет сетки. Вокруг первичных складок располагаются крупные пузырчатые клетки со светлой цитоплазмой. В этом возрасте начинают формироваться вторичные складки и в крупных (первичных) ячеях появляются (вторичные) мелкие ячеи. Ростковый слой представлен одним рядом призматических клеток. Мышечная оболочка уплотняется и утолщается за счет продольного слоя, серозная оболочка остается без изменений.
У 120-дневных плодов уже сформированы крупные и мелкие складки слизистой оболочки, появляется крупнопетлистая сеть, в ячеях последней – мелкие ячеи. Поверхностные клетки эпителия имеют светлые ядра. Мышечная оболочка утолщается за счет циркулярного слоя. В слоях мышечной оболочки появляются сосудистые сети, ячеи которых вытянуты вдоль мышечных волокон. Серозная оболочка, по сравнению с плодами 105 дней утолщается.
В возрасте 135 дней поверхностные пузырчатые клетки становятся плоскими и по размерам уменьшаются, базальные клетки расположены в один ряд и имеют форму призмы. Соединительнотканная основа уплотняется и в ней появляется разветвленная сосудистая сеть, толщина мышечной оболочки возрастает, при этом, толщина циркулярного и продольного слоев становится одинаковой. Серозная оболочка остается без изменения.
У новорожденных козлят слизистая оболочка сетки имеет ячеистый вид, пузырчатые клетки эпителия уменьшаются в размерах и их ядра занимают 50 % площади цитоплазмы. Ростковые клетки расположены в один ряд. Все структуры стенки, по сравнению с плодами 135 дней, остаются без изменения, лишь утолщается мышечная, серозная оболочки.
У коз в возрасте одного года первичные и вторичные складки слизистой оболочки покрыты плоским эпителием, в их соединительнотканную основу со стороны циркулярного слоя проходят пучки мышечной ткани. Мышечная оболочка представлена сформированными волокнами мышечных клеток, окруженных прослойками соединительной ткани и кровеносными сосудами. Серозная оболочка существенных изменений не претерпевает.
Гистогенез стенки книжки
Стенка книжки плода 60 дней представлена слизистой, мышечной и серозной оболочками. Слизистая оболочка образует большие и средние листочки, а между ними появляются два малых листочка. Чередование их следующее: между двумя большими листочками располагаются один средний и два малых листочка.
Эпителиальное поле листочков представлено ростковым слоем, состоящим из одного ряда клеток цилиндрической формы с апикально расположенными ядрами, а у основания листочков – 3–4 ряда. Крупные клетки со светлой цитоплазмой, располагаются в 1–2 слоя. Соединительная и мышечная ткани образуют основу листочков, в первой находятся крупные, овальные по форме фибробласты, во второй – миобласты. Серозная оболочка тоньше мышечной.
Стенка 75-дневных плодов более усложнена по строению. Ростковые клетки базального слоя листочков лежат в два ряда, а у их основания находятся крупные клетки со светлой цитоплазмой. Они увеличены в размерах и располагаются многослойно. Появляются самые малые листочки с примитивной мышечной оболочкой, разделяющейся на наружный – продольный и внутренний циркулярный слои. Серозная оболочка развита и содержит сеть кровеносных сосудов.
Стенка книжки 90-дневных плодов структурно оформлена. Крупные клетки со светлой цитоплазмой уменьшаются в размерах, четко выделяется ростковый слой, мышечная оболочка утолщается вследствие утолщения циркулярного слоя.Серозная оболочка покрыты мезотелием.
У 105-дневных плодов клетки со светлой цитоплазмой сдвигаются поверхностнее и уплощаются, ростковые клетки располагаются в один ряд. На листках появляются зачатки конических механических сосочков без роговых чехликов. Мышечная оболочка продолжает утолщаться, особенно, циркулярный ее слой. Он становится толще продольного почти в три раза. Серозная оболочка, по сравнению с 90-дневными плодами, уплотняется, становится тоньше.
У 120-дневных плодов клетки со светлой цитоплазмой становятся плоскими и расположены поверхностно, ростковые клетки лежат в один ряд. Мышечная оболочка продолжает утолщаться. Ее циркулярный слой толще продольного становится почти в пять раз. Из этого следует, что к рождению стенка книжки и листочки структурно соответствуют зрелым козам, но морфологически они до конца еще не оформлены.
Слизистая оболочка книжки и ее листки снаружи покрыта защитным слоем, который красится оксифильно, продолжает возрастать толщина мышечной оболочки стенки и ее листков. Циркулярный слой мышечной оболочки толще продольного почти в два раза. На конических механических сосочках появляются неоформленные до конца роговые чехлики.
Листочки книжки, 3–12-месячных козлят свободно располагаются в ее пространстве. Их эпителий покрыт защитным слоем, сформирован крупными клетками со светлой цитоплазмой, окрашивающиеся в оранжевый цвет. Циркулярный слой мышечной оболочки толще продольного в 1,5 раза, а сама оболочка увеличилась в толщине почти в 2 раза по сравнению с новорожденными козлятами. Серозная оболочка без изменений.
Гистогенез стенки сычуга
Стенка донной части сычуга 60-дневных плодов представлена слизистой, мышечной и серозными оболочками. Слизистая оболочка к этому времени имеет складки и желудочные ямки (крипты), которые в будущем преобразуются в донные железы сычуга. Они представляют впячивания многорядного эпителия в подслизистый слой оболочки и отличаются друг от друга разной степенью зрелости, одни находятся в стадии закладки, другие внедрились в подслизистую соединительную ткань в виде тяжей. Эпителий слизистой оболочки сычуга представлен многорядно расположенными цилиндрическими клетками с крупными шарообразными ядрами, сдвинутыми к апекальному полюсу. В соединительной ткани оболочки хорошо выражена волокнистость, в которой клеточные элементы концентрируются ближе к базальной мембране. В этом возрасте плодов слизистая оболочка толще мышечной в три и более раза.
Мышечная оболочка подразделяется на внутренний – циркулярный и наружный – продольный слои, последний очень слабо развит и между ними располагаются кровеносные капилляры. Серозная оболочка представлена слабо дифференцированными мезенхимными клетками и покрыта мезотелием.
У 75-дневных плодов стенка сычуга усложняется по строению. Эпителиальные клетки желудочковых ямок располагаются в один слой. В клетках, находящихся в области устья ямок, ядра сдвигаются к базальному полюсу, в остальных лежат в центре цитоплазмы. Область дна желудочковой ямки преобразуется в альвеолу, в стенке которой появляются одиночные обкладочные клетки со светлой цитоплазмой. В этом возрасте они увеличиваются в длину, просвет выводных протоков будущей железы зауживается, а соединительнотканные перегородки, разделяющие альвеолы уменьшаются в толщине. В подслизистом слое слизистой оболочки расположена разветвленная сеть кровеносных сосудов. Серозная оболочка без изменений.
Мышечная оболочка, по сравнению с 60-дневными плодами, утолщается за счет внутреннего циркулярного слоя. В ней часто встречается миобластические элементы, в этом возрасте продольный мышечный слой выражен слабее.
У 90-дневных плодов увеличивается складчатость слизистой оболочки сычуга. Железы глубоко располагаются в соединительной ткани, достигая мышечного слоя. Подслизистый слой развит лучше, толстый, содержит большое количество соединительной ткани. Мышечная оболочка в 1,5 раза толще, по сравнению с 75-дневными плодами. В этом возрасте, особенно, происходит увеличение продольного мышечного слоя. Серозная оболочка без изменений.
В возрасте плодов 105 дней в слизистой оболочке сычуга происходит уплотнение желез, лишь желудочковые ямки (крипты) остаются воронкообразно расширенными. Мышечным слоем слизистая оболочка подразделяется на собственный и подслизистый слои, между которыми располагаются кровеносные сети. Мышечная оболочка стенки сычуга утолщена, по сравнению с 90-дневными плодами, до двух раз, особенно, утолщается продольный слой, а между ее слоями развита сеть кровеносных сосудов.
В возрасте плодов 120 дней донные железы сычуга внешне сформированы, в желудочковые ямки открывается от двух до трех выводных протоков желез. Стенка концевого отдела железы покрыта кубическим эпителием, ядра их имеют полулунную форму
и располагаются на базальном полюсе клеток. В этом возрасте истончен подслизистый слой в 1,5 раза. Мышечная оболочка в целом утолщается, но цилиндрический слой толще продольного в два раза.
В возрасте плодов 135 дней происходит усложнение строения донной части стенки сычуга, увеличивается число желез, в железах –
количество главных и добавочных клеток, в подслизистом слое достоверно увеличивается количество тучных клеток. Утолщается в целом мышечная оболочка, причем возрастает толщина как циркулярного, так и продольного слоев. Из этого следует, что донная часть сычуга плодов 135 дней морфологически и функционально подготовлена к приему, перевариванию пищи после рождения.
У новорожденных козлят происходит удлинение складок слизистой оболочки сычуга. Увеличивается глубина, плотность и количество залегания донных желез. В концевых отделах их возрастает число обкладочных, но количество главных и добавочных клеток остается без изменения. Возрастает толщина мышечной оболочки. Брюшина остается без изменения.
У молодняка коз в возрасте 6 месяцев усложняется строение желез, в стенке донной части сычуга появляются лимфатические фолликулы, облегающие железы. Утолщается мышечная оболочка. Между ее слоями развита кровеносная сеть.
В возрасте коз 12 месяцев вершина складок слизистой оболочки раздваивается, что увеличивает площадь растяжения при наполнении сычуга кормами и их перевариваемую способность. Донные железы усложняются по строению. Они имеют укороченный концевой отдел и удлиненный выводной проток, открывающийся в желудочковые ямки. Лимфоидные образования в слизистой оболочке, разрастаясь, заполняют пространства между донными железами. Местами они выступают в просвет сычуга. Мышечный слой слизистой оболочки имеет равномерную толщину и располагается непосредственно под базальной мембраной. Мышечная оболочка стенки донной части сычуга разделена четко на слои и мышечные пучки. Ее циркулярный слой толще продольного слоя в 1,5 раза.
Из данного анализа следует, что стенки преджелудочков новорожденных козлят структурно и функционально подготовлены к перевариванию пищи после рождения, однако с изменением рода пищи (молозиво, молоко, грубый корм) меняется структура донной части сычуга.
Развитие тонкого кишечника
У мелкого рогатого скота образование петли первичной кишки просходит на 22 день эмбрионального развития (Л.В. Давлетова, 1974), путем оттягивания изгиба вниз желточным протоком к пупочному отверстию (В.Н. Жеденов, 1965). С появлением на 27 день развития выроста (будущая слепая кишка), петля подразделяется на восходящее и нисходещее колено. На 40 день развития нисходящее колено петли преобразуется в тонкий отдел кишечника, а восходя-
щее – в толстый. При этом весь кишечник интенсивно растет в длину.
У 25-дневных зародышей первичная кишка выстлана многорядным эпителием, 30-дневных предплодов в стенке кишки уже различимы три оболочки, а в эпителии увеличивается число ядер. Гистохимическими методами исследования в клетках установлено наличие гликогена, гликопротеидов, нуклеиновых кислот, АТФ и др. (Л.Л. Ошляк, 1986).
Известно, что для питания зародыша, раннего предплода большое значение имеют углеводы, которые они получают из маточного молочка, а также из клеток провизорных органов, особенно амниона. Этим объясняется наличие гликогена в клетках эпителия пищеварительной системы 30-дневных предплодов.
Начиная с 35 дней, в стенке кишки развивается циркулярный, а затем – продольный мышечные слои, появляются межмышечное нервное сплетение и ганглии, эпителий слизистой оболочки переходит в каемчатый. В 40 дней – в 12-перстной кишке предплодов появляется первичная складчатость, кишечные ворсинки, железы, в 45 дней эти процессы проходят в тощей, а у плодов 60 суток – подвздошной кишке.
У поздних и ранних предплодов энергично растет в длину тонкий кишечник, что приводит к образованию петель, складчатости и ворсинок слизистой оболочки и, как следствие, увеличевается его всасывающая поверхность.
К концу предплодного периода развития оболочки стенки кишечного тракта сформированы. Основа слизистой оболочки представлена эмбриональной соединительной тканью, а эпителиальное поле – многорядным и многослойным эпителием, мышечная оболочка подразделяется на циркулярный и продольные слои.
С формированием плаценты (27 дней) возникает гемотрофное питание предплода и плода. В итоге возникает обмен веществ между
кровотоком матери и плода, что приводит к интенсивному развитию пищеварительной системы. В связи с чем, в преджелудках поздних предплодов накапливается меконий, а дифференцировка слизистой оболочки приводит к возникновению железистого и всасывающего эпителия.
У ранних плодов вначале образуются ворсинки, крипты, затем железы в 12-перстной кишке, позже – в тощей и, в последнюю очередь – общекишечные железы в подвздошной кишке. Эпителиальные клетки слизистой оболочки образуют каемку, а среди каемчатого эпителия появляются бокаловидные клетки. Однако, в этом периоде, в клетках стенки тонкого кишечника сохраняется наибольшая концентрация гликогена, а в содержимом просвета – белков.
В позднеплодном периоде развития интенсивность тонкого отдела и обменных процессов у плода возрастает, усиливается транспорт белков кровью в сторону просвета кишечника. Об этом свидетельствует появившаяся вакуолизация эпителия: в 12-перстной кишке – 60, в тощей – 75 и подвздошной – 105-дневных плодов. В верхушке ворсинок под эпителием видны обширные полости. С этого времени отмечается высокая концентрация белка в слизистой и мышечной оболочках (Л.Л. Ошляк, 1986), что сопровождается инфильтрацией их лимфоцитами, и с этого момента начинает накапливаться меланин в кишечнике.
Дифференциация мышечной оболочки кишки на циркулярный и продольный слои заканчивается у 105-дневных, в подвздошной – 120-дневных плодов. В слизистой оболочке, соответственно, появляются лимфатические узелки, у 135-дневных плодов они достигают высокой дифференциации, к рождению часть их смещается под эпителий, однако функционально они еще не готовы, к защите организма.
Таким образом, к рождению козлят структура стенок тонкого кишечника достигает высокой степени развития, соответствует структуре кишечника коз после рождения и подготовлена в морфо-функциональном плане к смене условий существования и питания козлят в первые дни после рождения.
С рождением козлят перестает функционировать плацентарное кровообращение, ослабляется межуточный обмен веществ, все это отражается на обменных функциях пищеварительной системы, что приводит к снижению проницаемости слизистой оболочки кишечного тракта. После рождения через 24–36 часов между эпителиальным полем и соединительной тканью подслизистого слоя формируется базальная мембрана, эпителиальные клетки приобретают ворсинки, исчезает межклеточная пористость, пинацитозные вакуоли, границы клеток становятся гомогенными. Таким образом, перекрывается путь поступления гамма-глобулинов и лимфацитов из молозива матери в кровь новорожденных.
Известно, иммунологической функцией обладает гамма-глобулины. При исследовании плодов установлено, что с возрастом в сыворотке увеличивается обще количество белка за счет альбуминов и в меньшей степени за счет альфа, бета-глобулинов. Гамма-глобулины у поздних плодов и новорожденных отсутствуют (Л.П. Давлетова, 1974) и др.
Одной из причин отсутствия гамма-глобулина в крови новорожденных козлят является высокая молекулярная масса, что препятствует прохождению белка из крови матери через плаценту в кровь плода. В целом передача антител от матери к плоду может осуществляться, по мнению Л.В. Давлетовой (1974), только из молозива после рождения через стенку кишечника. Установлено, что гамма-глобулины захватываются коническими клетками 12-перстной, подвздошной кишок и посредством пинацитоза переносятся в вакуоли между ворсинками, а оттуда они попадают в лимфу и кровь плода, но спустя 18 часов после рождения, клетки не адсорбируют белки и в них, по этой причине, отсутсвуют уже вакуоли.
С. Георгиев (1969), П.А. Емельяненко (1987) признают адсорбцию белков до 48 часов после рождения. Поэтому, количество гамма-глобулина в крови козлят может возрастать до 48 часов после рождения. Однако, если молозиво не давать новорожденному в течение 36 часов, то гамма-глобулин вообще не обнаруживается в их крови (Н.Н. Гриценко, 1968).
Большое значение имеет в козоводстве получение молозива новорожденными в первые часы жизни после рождения, так как через 36–48 часов после рождения молозиво становится молоком, а в молоке, отсутствуют антитела. Отсюда этот период имеет большое значение в сохранности новорожденных козлят.
В дальнейшем происходит совершенствование структур стенки и, в первую очередь, железистого аппарата тонкого кишечника. Увеличивается число ворсинок, вторичных складок, крипт и дуоденальных желез, что усиливает пищеварительную функцию и увеличивает всасывающую поверхность кишечника. В экскрете желез равномерно увеличивается количество гликопротеидов, достигая максимума у коз в возрасте 6–9 месяцев. У новорожденного козленка длина тонкого кишечника достигает 7 м, в годичном возрасте – 16,9 м и устанавливается в трехлетнем возрасте. Затем до семи лет длина возрастает, но темп ее роста снижается. Об этом свидетельствует абсолютный прирост и относительный рост длины кишечника (табл. 72).
Таблица 72
Динамика роста тонкого отдела кишечника, см
|
Возраст |
Кол-во |
Длина |
Абсолютный |
Относительный рост, раз |
|||||||||
|
12-перстной |
Тощей |
Подвздошной |
Всего кишечника |
12-перстной |
Тощей |
Подвздошной |
Всего кишечника |
12-перстной |
Тощей |
Подвздошной |
Всего кишека |
||
|
Новорожденный |
5 |
34,0 |
639, |
11,0 |
693 |
– |
– |
– |
– |
– |
– |
– |
– |
|
1 мес. |
3 |
43,0 |
1055 |
13,5 |
1132 |
9,0 |
416,5 |
2500 |
439 |
1,26 |
1,65 |
1,20 |
1,63 |
|
6 |
4 |
58,0 |
1360 |
17,0 |
1435 |
15,0 |
380 |
3500 |
303 |
1,34 |
1,28 |
1,26 |
1,27 |
|
12 |
3 |
88,0 |
1580 |
19,0 |
1687 |
30,0 |
220 |
2,0 |
296 |
1,51 |
1,16 |
1,12 |
1,21 |
|
36 |
3 |
92,0 |
1728 |
21,0 |
1841 |
4,0 |
148 |
2,0 |
149 |
1,05 |
1,09 |
0,97 |
1,09 |
|
72 |
3 |
93,0 |
2132 |
23,0 |
2248 |
1,0 |
404 |
2,0 |
402 |
1,01 |
1,23 |
1,02 |
1,21 |
|
По В.Н. Жеденову (1965) |
50,0 |
2620 |
30,0 |
2700 |
– |
852 |
– |
820 |
– |
– |
– |
– |
|
Из данного анализа следует, со сменой рода пищи (молозиво, молоко, грубый корм) изменяется структура и морфология стенки кишки, увеличивается объем и ее всасывающая поверхность. В связи с этим нарушение технологии содержания, ухода, питания обуславливает понижение иммунологических функций органов защиты, что приводит к повышенной восприимчивости коз к инфекционным и другим заболеваниям и падежу. Поэтому очень важно чтобы новорожденные козлята как можно раньше принимали молозиво, а с ним иммунные тела.
Развитие печени
Источником образования печени является первичная кишка, из которой на 19 день эмбрионального развития, после зачатка желудка, появляется выпячивание, впоследствии превратится в печень (О.Б. Шумкина, 1960). По наблюдению В.К. Бирих, Г.М. Удовина (1972), Л.В. Давлетовой (1974), О.В. Волковой, М.И. Пекарского (1976) из всех органов пищеварения предплодов наиболее активно развивается печень. Она заполняет почти всю их брюшную полость. Желудок и кишечник в этом возрасте располагается на внутренней поверхности печени, между ее долями.
В паренхиме печени предплодов энергично идут процессы кроветворения и дифференциации, возрастает число гепатоцитов. В предплодном и раннем плодном периодах развития печеночные клетки (А.Г. Термелева, 1970) синтезируют гликоген. Высказано предположение, что по мере формирования органа гликоген идет на развитие печени и гемопоэза. В начале раннего плодного периода развития печень достигает неибольшей относительной массы, к концу – итенсивность роста ее снижается и по объему она все больше уступает развитию жлудка и кишечника. В начале позднего плодного периода процессы гемопоэза затухают, а на смену им приходит функция пищеварения.
Уже в раннем плодном периоде развития структура печени приобретает пластинчатую форму строения (Л.В. Давлетова, 1974), начинает вырабатывать липазу, желчные кислоты и выделять желчь. В печени поздних плодов появляется дольчатость, а в них – кровеносные, желчные капилляры. Гепатоциты ориентированы радиально относительно к центральной вене дольки. В это время, в клетках интенсивно происходит накопление гликогена. Из этого следует, что к концу плодного периода развития печень достигает морфологической и функциональной зрелости. Из этого следует, что в предплодном, раннеплодном периодах развития печеньь является основным органом кроветворения, поэтому имела значительные размеры и гистологическое строение близкое к органам гемопоэза. К концу плодного периода, в связи с началом пищеварения и развитием скелета, функция гемопоэза, с возрастающей активостью переходит к костному мозгу, что сказывается на росте размеров, на морфологической и функциональной зрелости печени. Одновременно, замедление роста печени во второй половине плодного развития, резко ускоряет рост скелета, собенно, периферического (В.К. Бирих, Г.М. Удовин, 1972).
Масса печени с возрастом плодов и коз возрастает неравномерно (табл. 73) и устанавливается в три года, затем до пяти лет увеличивается, но с меньшей активностью.
Наибльший прирст массы печени отмечается в три, 12 и 18 месяцев, в относительных величинах – в один, три, 12 месяцев, затем темп роста остается на одном уровне. Из этого следует, что масса печени коз устанавливается в возрасте 12–18 месяцев.
Таблица 73
Динамика роста массы печени коз
|
№ п/п |
Возраст |
Количество |
Абсолютная |
Прирост, |
Относительный |
|
1 |
120 дней |
3 |
49,0 |
- |
- |
|
2 |
Новорожденные |
3 |
54,0 |
5,0 |
1,1 |
|
3 |
1 мес. |
3 |
98,0 |
44,0 |
1,8 |
|
4 |
3 |
2 |
213,5 |
133,0 |
2,4 |
|
5 |
6 |
3 |
289,6 |
58,1 |
1,3 |
|
6 |
12 |
2 |
460,4 |
170,8 |
1,6 |
|
7 |
18 |
2 |
608,0 |
147,6 |
1,3 |
|
8 |
3 года |
3 |
674,5 |
65,5 |
1,1 |
|
9 |
5 лет |
3 |
783,5 |
109,0 |
1,1 |
|
Итого: |
24 |
Развитие поджелудочной железы
Поджелудочная железа закладывается из первичной кишки, после зачатка печени, на 22 день эмбрионального развития (О.Б. Шумкина, 1960), а печени – на 19 день. Как показали исследования Л.Т. Капраловой (1971) железа предплодов резко увеличивает массу, в ее паренхиме появляюся первичные протоки, клетки которых дают начало образованию ацинусов и панкреатических островков. Первые островки появляются на 30, а ацинусы на 45 день плодного периода развития, т.е. несколько позже закладки островков. Из этого следует, что островки уже играют важную роль в развитии предплодов.
Поджелудочная железа уже ранних плодов проявляет протеолитическую и амилолитическую активности. В этом возрасте в ацинусах накапливаются гранулы зимогена. Железа поздних плодов, в возрасте 90–105 дней, принимает активное участие в углеводном обмене, в панкреатических островках появляются «А» и «В» клетки. Появление «В» клеток совпадает с началом депонирования гликогена в печени, железа начинает выделять липазу. Поэтому к протеолитической, амилолитической функциям добавляется липолитическая активность. К рождеию железа четко подразделяется на экзокринную и эндокринную части.
У новорожденных экзокринная часть достигает 46,7 %, а у взрослых 96–98 % массы поджелудочной железы, а эндокринная значитльно меньше – 2–4 % (Л.В. Давлетова, 1971). Между ацинусами располагаются скопления клеток, называемые панкреатическими островками, размером от 20 до 360 мкм. Островки состоят из альфа –
клеток, расположенных по периферии и синтезирующие глюкагон, более многочисленных – бета-клеток, вырабатывающих инсулин, и дельта – клеток, образующие соматостатин, подобный соматостатину гипоталамуса (Дж. Бакл, 1986).
Железа новрожденных светло-красного цвета, располагается в дубликатуре S-образного изгиба 12-перствной кишки, частично в воротах печени. В этом возрасте различают тело, левую, правую доли и хвостатый отросток. Левая доля лежит ниже задней полой вены, выходит на внутреннюю стенку рубца, срастаясь с ней, достигает медиального края селезенки. Тело и правая доля лежат в брыжейке S-образного изгиба 12-перствной кишки. Хвостатый отросток двойной, с дорсальной и вентральной поверхностей охватывает заднюю полую вену, вытягиваясь назад до 11 см (Б.П. Шевченко, М.Ю. Маховых, 1999). Проток поджелудочной железы открывается вместе с желчным в 12-перстную кишку на расстоянии до 10 см от пилоуса сычуга.
Масса поджелудочнй железы с возрастом коз увеличивается неравномерно, наиболее интенсивно она растет у плодов до 120 дней, к рождению несколько замедляется, затем к месячному возрасту после рождения увеличивается, но незначительно, осбенно, активно масса железы возрастает в три и в 12 месяцев. К трем годам масса железы, по сравнению с 12-месячными козами, практически удваивается (табл. 74). К пяти годам масса железы умеьшается на 5,1 г. Из данного анализа следует, что с возраста пяти лет наступают инволютивные процессы железы.
Таблица 74
Динамика роста массы поджелудочой железы коз
|
№ п/п |
Возраст |
Кол-во |
Абсолютная масса, г |
Прирост, г |
Относительный рост, раз. |
|
1 |
120 дней |
3 |
0,27 |
– |
– |
|
2 |
Новорожденные |
3 |
1,26 |
0,99 |
4,70 |
|
3 |
1 мес. |
3 |
1,70 |
0,44 |
1,35 |
|
4 |
3 |
3 |
2,61 |
0,91 |
1,54 |
|
5 |
6 |
3 |
9,35 |
6,74 |
3,58 |
|
6 |
12 |
3 |
12,71 |
3,36 |
1,36 |
|
7 |
18 |
2 |
22,6 |
9,89 |
1,78 |
|
8 |
3 года |
3 |
49,8 |
27,20 |
2,20 |
|
9 |
5 лет |
3 |
44,7 |
–5,1 |
1,10 |
|
Итого: |
25 |
Наступает |
Развитие толстого кишечника
Толстый отдел кишечника развивается из восходящего колена первичной кишки, на 37–39 день плодного периода развития слепой отросток, лежащий на границе нисходящего и восходящего колена, преобазутся в слепую кишку. При этом, Э.М. Бикчентаев (1979) указывает, что абсолютная скорость роста кишечника от возраста к возрасту на всем потяжении плоднго развития повышается. За исследованный период (60–75 дней) абсолютная скорость роста тонкого кишечника была значительно вьше толстого. Так, среднесуточный прирост тонкого кишечника составил 0,6 г, а толстого лишь – 0,2 г. (табл. 75). К концу первой половины плодного развития разница в диспропорции сокращается. Однако, за 15 суток до рождения, тонкий отдел кишечника вновь преобладает в росте над толстым.
Тенденция опережающего роста тонкого кишечника по отношеию к толстому сохраняется и в постнатальном периоде развития (табл. 76) и связан с тем, что в тонкой кишке продолжается в основном химическое переваривание пищи и частично всасывание, а в толстом – всасывание, формирование каловых масс и эвакуация их наружу.
Наиболее рельефно опережающий рост тонкой кишки выражен до возраста козлят одного месяца (6,0–6,3 раза), затем темп его роста постепенно понижается до трех лет. Если рассматривать пищеварительную систему с точки отношения её к живой массе, то с возрастом коз на один метр тонкой и толстой кишок в отдельности в новорожденном возрасте приходится, соответственно, 0,46 и 2,8 кг, а в три года – 2,14 и 7,21 кг, то есть нагрузка на толстый кишечник выше в 3,8 раза, чем на тонкий (табл. 76).
Таблица 75
Динамика скорости роста массы кишечника, г
(по Э.М. Бикчентаеву)
|
Возраст плодов, дн. |
Плоды |
Кишечника |
Тонкого |
Толстого |
||||
|
Абсолютный рост |
Среднесуточ. прирост |
Абсолютный рост |
Среднесуточный прирост |
Абсолютный рост |
Среднесуточный прирост |
Абсолютный рост |
Среднесуточный прирост |
|
|
60–75 |
99,9 |
6,66 |
0,88 |
0,06 |
0,68 |
0,05 |
0,20 |
0,01 |
|
75–90 |
236,2 |
15,75 |
1,65 |
0,11 |
1,23 |
0,08 |
0,42 |
0,03 |
|
90–105 |
421,7 |
28,12 |
3,72 |
0,25 |
2,61 |
0,18 |
1,11 |
0,07 |
|
120–135 |
676,5 |
45,10 |
16,40 |
1,09 |
12,38 |
0,82 |
4,02 |
0,27 |
|
135–новорожденные |
833,8 |
55,58 |
42,70 |
2,85 |
32,43 |
2,16 |
10,27 |
0,69 |
|
60?новорожденные |
2941,1 |
32,68 |
72,44 |
0,81 |
54,29 |
0,61 |
18,15 |
0,20 |
Из анализа возрастных изменений пищеварительной системы коз видно, что рост массы и линейных размеров кишечника в плодном и в постнатальном периодах развития происходит нерав-
номерно.
Абсолюбтная скорость роста кишечника на протяжении онтогенеза повышается, достигая максимума перед рождением и до одного месяца после рождения. Основной прирост массы кишечника происходит во второй половине плодного и до года постнатального развития.
Таблица 76
Рост отделов кишечника относительно друг друга и к массе коз
(по Э.М. Бикчентаеву)
|
Возраст |
Масса, кг |
Тонкий отдел, см |
Толстый отдел, см |
Относительный рост, раз |
Масса тела на 1 м тонкого отдела |
Масса тела на 1 м толстого отдела |
|
Новорожденные |
3,2 |
693,0 |
115,0 |
6,03 |
0,46 |
2,80 |
|
1 мес |
8,5 |
1055,0 |
167,0 |
6,30 |
0,80 |
5,10 |
|
6 |
21,2 |
1360,0 |
379,0 |
3,60 |
1,56 |
5,60 |
|
12 |
24,6 |
1580,0 |
487,0 |
3,24 |
1,56 |
5,05 |
|
36 |
37,0 |
1728,0 |
513,0 |
3,37 |
2,14 |
7,21 |
|
72 |
46,0 |
2244,0 |
590,0 |
3,80 |
2,05 |
7,80 |
Таблица 77
Динамика роста толстого отдела кишечника
|
Возраст |
Кол-во |
Длина |
Абсолютный |
Относительный рост, раз |
|||||||||
|
слепая |
ободочная |
прямая |
всего |
слепая |
ободочная |
прямая |
всего |
слепая |
ободочная |
прямая |
всего |
||
|
Новорожд. |
5 |
10 |
93 |
12 |
115 |
– |
– |
– |
– |
– |
– |
– |
– |
|
1 мес. |
3 |
27 |
119 |
17 |
167 |
21,0 |
26,0 |
5,0 |
52 |
2,7 |
1,3 |
1,41 |
1,45 |
|
6 |
4 |
36 |
320 |
23 |
379 |
5,0 |
201 |
6,0 |
212 |
1,2 |
2,7 |
1,4 |
2,3 |
|
12 |
3 |
43 |
420 |
24 |
487 |
7,0 |
100 |
1,0 |
108 |
1,2 |
1,3 |
1,05 |
1,3 |
|
36 |
3 |
44 |
443 |
26 |
513 |
1,0 |
23 |
2,0 |
26 |
1,02 |
1,1 |
1,08 |
1,05 |
|
72 |
3 |
45 |
520 |
25 |
520 |
1,0 |
77 |
– |
77 |
1,02 |
1,17 |
– |
1,15 |
|
Взрослых по В.Н. Жеденову (1965) |
30 |
450 |
20 |
500 |
– |
– |
– |
– |
– |
– |
– |
– |
|
Особого внимания заслуживает рост кишечника перед рождением и до одного месяца после рождения, когда рост массы тела козлят ежемесячно более чем удваивается. Цитируемые данные указывают на чрезвычайную важность уровня кормления, содержания беременных и подсосных козоматок для получения и выращивания жизнеспосбного молодняка.
У новорожденных козлят длина слепой кишки достигает 10,0 см, ободочной – 93,0 см, прямой – 12,0 см (табл. 77). Наиболе интенсивно тостый отдел кишечника увеличивается в длину до одного месяца после рождения, затем темп роста уменьшается до шести месяцев, хотя остается высоким, и устанавливается в 12 месяцев.