
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

6.5. Фотосинтетическая деятельность
Исследованиями А.А. Ничипоровича (1966) и А.Я. Бакалдина (1973) установлено, что урожай сухого вещества сельскохозяйственных культур на 80–90 % создается в результате фотосинтеза, который, в первую очередь, зависит от размеров ассимиляционной поверхности, высоты и густоты стояния растений и ряда других факторов. Все остальные процессы питания растений, в частности водное и минеральное, эффективны в той степени, когда они обеспечивают и поддерживают оптимальную деятельность фотосинтетического аппарата.
Нарастание сухой массы считается одним из главных показателей фотосинтетической деятельности растений. Суточный прирост сухого вещества значительно изменяется и может достигать до 300 кг/га в период интенсивного роста растений (А.А. Ничипорович и др., 1961).
Потенциальная биологическая продуктивность полевых культур зависит от величины поступающей к ним солнечной энергии и от особенностей её расхода. Помимо продуцирования кислорода, фотосинтез приобретает всё большее значение как основа воспроизводства продовольствия. Поэтому, создавая искусственные ценозы на полях, человек при помощи агротехники и лучистой энергии солнца обеспечивает себя продуктами питания. Первичные продукты фотосинтеза, участвуя в реакции вторичного метаболизма, образуют весь спектр органических соединений растительной клетки (А.Т. Мокроносов, В.Ф. Гавриленко, Т.В. Жигалова, 2006).
Высокая продуктивность сельскохозяйственных культур определяется динамически оптимальным соотношением отдельных элементов фотосинтеза. К основным из них относят: размер ассимиляционного аппарата, фотосинтетический потенциал, интенсивность и продуктивность фотосинтеза (Т.М. Русакова, 1974; J.M. Anderson, 1983; В.М. Важов, 2012).
Известно, что годовая масса органического вещества, создаваемого фотосинтезом, с избытком перекрывает то количество продовольствия, которое необходимо населению планеты. Поэтому одним из путей повышения эффективности земледелия является создание посевов с хорошей структурой пространственного расположения листьев.
Листовая поверхность является главным рабочим органом зелёных растений, а размер урожая почти полностью определяется её величиной. Нарастание листовой поверхности и величина ассимиляционного аппарата культурных растений зависит от агротехнических условий (А.Н. Бегишев,1953; Vong Hguyen Quoc, Murata Yoshio, 1978) .
По вопросу об оптимальной площади листовой поверхности сельскохозяйственных культур нет единого мнения. А.А. Ничипорович (1959) считает, что листовая поверхность должна составлять до 50 тыс. м2/га, а по мнению А.Н. Бегишева (1953) этот показатель равен примерно 100 тыс. м2/га.
В исследованиях А.Д. Гончарова (2008) в Новосибирской области бороздковые посевы формировали лучшую листовую поверхность гречихи, чем сплошные. С возрастом растений преимущество бороздковых посевов было очевидным. В фазу бутонизации площадь листьев на 1 га бороздковых посевов была больше рядовых на 14,5 %, а в фазу цветения – на 37,4 %. В изменении площади листьев по годам исследований каких-либо закономерностей не отмечено как по каждому способу возделывания, так и по сравнению их между собой. В различиях между изучаемыми способами посева по другим показателям фотосинтетической деятельности гречихи и их динамики в течение вегетационного периода в опытах А.Д. Гончарова (2008) проявились те же закономерности, что и в случае с площадью листьев. Так, фотосинтетический потенциал бороздковых посевов был больше соответствующего показателя сплошных посевов на 15 % за период бутонизация-цветение и на 30 % – за время цветение-созревание.
По мнению Н.Д. Кумсковой (2004), в условиях Дальнего Востока на листовую поверхность гречихи влияют сроки сева. В её опытах посевы 5 и 30 июня имели площадь листьев 22,5 и 23,5 тыс. м2 на 1 га, тогда как посевы 20 и 25 июня, соответственно, 32 и 31 тыс. м2. Гречиха давала хорошую вегетативную массу и высокие показатели фотосинтеза. В опыте Н.Д. Кумсковой (2004) при посеве с 5 по 30 июня получено от 25,1 до 37,3 т/га зелёной массы, максимальной она была при сроке сева 20 июня (табл. 56).
Таблица 56
Фотосинтетическая деятельность и урожайность вегетативной массы гречихи
на разных сроках сева (по Н.Д. Кумсковой, 2004)
|
Срок сева |
Площадь листьев (тыс. м2/га) |
Зеленая масса, т/га |
Сухая масса, т/га |
ЧПФ |
|
5VI |
22,5 |
25,1 |
5,4 |
3,6 |
|
15 VI |
28,0 |
33,2 |
6,3 |
4,6 |
|
20 VI |
32,0 |
37,3 |
7,2 |
8,0 |
|
25VI |
31,0 |
33,6 |
6,9 |
7,8 |
|
30V1 |
23,5 |
27,6 |
6,5 |
6,2 |
Примечание. ЧПФ – чистая продуктивность фотосинтеза за вегетацию, г/м2/сут.
По мнению Н.Д. Кумсковой (2004), влияние удобрений на площадь листьев, продуктивность фотосинтеза, прирост сухого вещества гречихи значительное (рис. 61).
Прирост зелёной и сухой массы гречихи находился в тесной зависимости от удобрений. На вариантах без их применения и при внесении азота прирост зелёной массы продолжался до уборки, что важно при выращивании гречихи на зелёное удобрение. Внесение фосфора, совместно с комплексным удобрением, обеспечивало накопление биомассы на 1–2-е декады раньше (табл. 57).
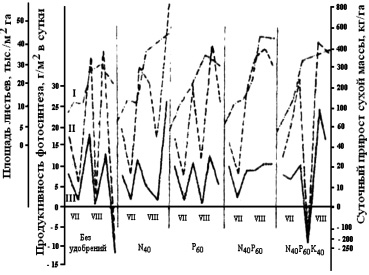
Рис. 61. Влияние удобрений на рост площади листьев, продуктивность фотосинтеза, прирост сухого вещества гречихи по декадам (по Н.Д. Кумсковой, 2004)
Таблица 57
Динамика нарастания зелёной и сухой массы гречихи в зависимости от удобрений (по Н.Д. Кумсковой, 2004), т/га
|
Удобрение |
10/VII |
20/VII |
30/VII |
10/VIII |
20/VIII |
Высота раст., см |
|
Зелёная масса |
||||||
|
Без удобрений |
4,39 |
9,12 |
25,2 |
28,7 |
29,7 |
98 |
|
N40 |
3,82 |
8,48 |
30,3 |
37,1 |
37,4 |
104 |
|
P60 |
4,22 |
10,8 |
26,0 |
36,5 |
35,5 |
94 |
|
N40P60K40 |
2,34 |
13,5 |
35,6 |
33,1 |
31,8 |
104 |
|
Сухая масса |
||||||
|
Без удобрений |
0,6 |
1,2 |
3,3 |
6,5 |
7,1 |
- |
|
N40 |
0,6 |
1,4 |
3,6 |
5,5 |
7,2 |
- |
|
P60 |
0,5 |
0,9 |
2,9 |
6,0 |
8,3 |
- |
|
N40P60K40 |
0,3 |
2,2 |
3,9 |
5,7 |
8,4 |
- |
Удобрения, в зависимости от влажности почвы, по-разному влияли на формирование листовой поверхности гречихи (Н.Д. Кумскова, 2004). При выпадении большого количества осадков в начальный период вегетации на варианте с азотом формировалась низкая площадь листьев – 21,7 тыс. м2 на 1 га. На делянках, удобренных только фосфором и при совместном внесении азота, фосфора и калия листовая поверхность была больше на 19,3 и 25,1 тыс. м2 на 1 га, по сравнению с вариантом N40, и на 8,4 и 14,2 тыс. м2 – по отношению к контролю.
Существенно изменялась листовая поверхность в зависимости от густоты посева (Н.Д. Кумскова, 2004). С увеличением нормы высева с 2 до 5 млн. всх. зёрен на 1 га площадь листьев на одном растении снизилась с 175 до 104 см2, а на 1 га увеличилась с 26 до 57 тыс. м2.
Т.М. Русакова (1974) также считает, что площадь листьев гречихи на высоком агрофоне может превышать показатели контроля в 2,0–2,5 раза.
С.У. Броваренко (1970), на основании многолетних исследований в Западной Сибири, пришёл к выводу о том, что лучшее развитие ассимиляционной поверхности листьев гречихи происходит на узкорядных посевах, в сравнении с широкорядными. Площадь листьев на узкорядных посевах достигает 44,4 тыс. м2 на 1 га, на рядовых – 29,8, а на широкорядных – только 19,6 тыс. м2 на 1 га, соответственно, и урожайность на последних ниже. Можно высказать предположение, что урожайность на широкорядных посевах снижалась из-за угнетения культурных растений сорняками, так как известно, что рядовые, и особенно узкорядные посевы, успешно противодействуют засорённости.
С.И. Рак (1967) также считает, что площадь листьев гречихи при рядовом способе посева примерно в 1,5 раза больше в сравнении с листовой поверхностью на междурядьях 0,45 м.
По мнению К.А. Савицкого (1970), большое производственное значение имеют широкорядные посевы гречихи. На чистых от сорняков почвах они эффективнее сплошных рядовых. Листовая поверхность на широкорядном посеве в 1,3–1,4 раза больше, чем на сплошном. Это способствует повышению продуктивности фотосинтеза у растений широкорядного посева в фазе плодообразования в 1,5–1,9 раза.
Наши наблюдения подтверждают данные результаты. В зависимости от ширины междурядий площадь листьев возрастает в 1,4 раза в пользу разреженного посева, увеличение нормы высева также способствует росту ассимиляционной поверхности примерно в 1,5 раза.
Преимущество узкорядного способа посева перед обычным рядовым и широкорядным можно объяснить более слабым ростом, развитием и ветвлением растений в разреженных посевах, что наблюдается при наличии сорняков в междурядьях.
Гречиха интенсивно формирует листья в фазу всходов и бутонизации, к началу цветения их доля составляет 30–50 % от всей биомассы (Г.В. Копелькиевский, 1963). Способы посева и нормы высева значительно влияют на формирование площади листьев. Так, к периоду начала плодообразования размер листовой поверхности быстро увеличивается, но затем скорость данного процесса снижается.
Наши наблюдения показали, что в условиях лесостепи Алтайского края площадь листьев гречихи в фазу плодообразования составляет 48,9–69,8 тыс. м2/га (табл. 58).
Таблица 58
Площадь листьев и продуктивность фотосинтеза посевов гречихи
(средняя за 2010–2011 гг.)
|
Вариант |
Норма высева, млн. всх. зёрен на 1 га |
|||||
|
2,5 |
3,5 |
4,5 |
||||
|
площадь листьев, тыс. м2/га |
Фч. пр., |
площадь листьев, тыс. м2/га |
Фч. пр., |
площадь листьев, тыс. м2/га |
Фч. пр.,
г/м2 |
|
|
Рядовой (контроль) |
32,1 |
3,52 |
41,4 |
4,61 |
48,9 |
5,12 |
|
Широкоряд-ный (0,45 м) |
43,2 |
4,76 |
56,7 |
5,83 |
67,4 |
6,91 |
|
Широкоряд-ный (0,60 м) |
45,6 |
5,13 |
59,8 |
5,92 |
69,8 |
7,34 |
В связи с тем, что по урожайности зерна гречихи широкорядные посевы 0,60 м уступают таковым с междурядьями 0,45 м, можно предположить, что солнечная радиация и питательные вещества в разреженных посевах используются на создание вегетативной массы, а не зерна.
Таким образом, создавать оптимальную площадь листьев гречихи можно различными технологическими приёмами. Подбор лучших агротехнических условий, обеспечивающих оптимальное развитие листовой поверхности, имеет важное практическое значение.
Главным показателем фотосинтетической деятельности растений является чистая продуктивность фотосинтеза. Связь её с урожаем самая непосредственная и при равенстве площади листьев урожай сухой биомассы растений прямо пропорционален чистой продуктивности фотосинтеза (Т.М. Русакова, 1974; Schilling Herbert, 1980; J.H. Golbeck, 1992).
Продуктивность фотосинтеза гречихи не остается постоянной в течение всего периода вегетации. Наибольшую величину она имеет в период бутонизации-цветения, то есть в период наибольшего прироста сухой массы растений (Т.М. Русакова, 1974). Чистая продуктивность фотосинтеза следует за уровнем минерального питания: наиболее высокие показатели её наблюдаются у растений на чернозёмной почве с внесением минеральных удобрений (7,44 г/м2 в сут. в среднем за вегетацию).
Внесение минеральных удобрений в опытах Т.М. Русаковой (1974) довольно результативно отразилось на величине чистой продуктивности фотосинтеза. Это можно объяснить тем, что хорошее минеральное и, прежде всего, азотное питание активизирует фотосинтез деятельных, хорошо освещённых листьев верхних и средних ярусов, которые в сильной степени поддерживают активную жизнедеятельность листьев нижних ярусов. Последние, в условиях ослабленного света и затруднённого минерального питания, обычно быстро стареют и отмирают. Очевидно, поэтому на высоких фонах минерального питания образуются растения с большой площадью листьев и с высокими показателями чистой продуктивности фотосинтеза.
Анализ результатов определения чистой продуктивности фотосинтеза гречихи в условиях Новосибирской области говорит о том, что изменение этого показателя во времени и по способам возделывания культуры происходит по аналогии с изменением площади листьев и фотосинтетического потенциала. При этом, независимо от способа возделывания, чистая продуктивность фотосинтеза всегда была выше в благоприятные по увлажнению годы (А.Д. Гончаров, 2008).
По мнению Н.М. Городнего (1980), гречиха имеет лучшую, чем многие культуры способность к синтезу органических кислот (в мг на 1 г сухого вещества растения): гречиха – 7,01; нут – 5,08; люпин синий – 4,60; горчица – 4,55; горох – 3,02; овес – 2,88; кукуруза – 1,38. Всё это позволяет считать гречиху нетребовательной к почвам и приводит к тому, что её часто высевают на малоплодородной почве, где другие культуры почти не дают урожая.
По подсчетам А.А. Ничипоровича (1959, 1966), посевы культурных растений в процессе фотосинтеза способны образовывать до 8–10 г сухой биомассы на 1 м2 листовой поверхности, а потенциальная продуктивность может достигать 20–40 г/м2
в сутки. Этот показатель зависит от освещённости солнцем, притока питательных веществ и воды.
Гречиха имеет много точек роста, которые потребляют большое количество продуктов ассимиляции. Ассимиляция проходит при фотосинтезе, который напрямую зависит от величины листовой поверхности и интенсивности солнечного освещения. Величина листовой поверхности гречихи по отношению к количеству цветков на растении составляет всего 0,30–0,56 см2 на один цветок. Эта площадь слишком мала для полного обеспечения цветка пластическими веществами. Если период быстрого роста побегов совпадает со временем развития цветков и плодов, то при недостатке продуктов ассимиляции растущие верхушки стеблей и ветвей оказываются обеспеченными пищей лучше, чем большая часть репродуктивных органов (Н.М. Городний, 1980; J. Oppereer, 1985; S. Wojcik, 1991).
Листья гречихи не обладают высокой энергией усвоения углекислоты. При высокой температуре днём, даже при достаточной влагообеспеченности растений, листья подвядают. Фотосинтез ухудшается из-за быстрого роста вегетативных органов. В результате листья нижнего яруса сильно затеняются и освещаются рассеянным светом, существенно потерявшим физиологически активные лучи. Это снижает образование пластических питательных веществ, недостаток которых вызывает отмирание цветков и завязей (Vong Hguyen Quoc, 1977; И.Н. Елагин, 1984).
Гречиха – светолюбивое растение. Чем лучше освещены её листья, тем энергичнее и больше она усваивает из воздуха углекислоту, перерабатывает её в легкоусвояемые питательные вещества.
В условиях почвенной засухи рост гречихи прекращается, но развитие её продолжается, причем происходит оно быстрее, чем в нормальных условиях. В результате образуются карликовые растения, которые быстро отцветают и созревают, а урожай зерна резко снижается (Н.М. Городний, 1980).
По мнению М.Н. Городнего (1980), на загущенных посевах верхний ярус листьев гречихи поглощает только 60–70 % солнечной радиации. Средний и нижний ярусы получают третью часть и меньше общей энергии солнечного света. В среднем ярусе широкорядного посева проникающая радиация составляет 60–80 % общей падающей, а в нижнем – 50–70 %. Разреженный посев лучше использует световой режим для процессов фотосинтеза, по сравнению с загущенным.
Известно, что при затенении нижних ярусов листьев растений чистая продуктивность фотосинтеза начинает уменьшаться. Лучший по количеству и качеству урожай зерна гречихи можно получить только в посевах, обладающих достаточной по размерам площадью листьев – 40–45 тыс. м2/га при оптимальном ходе её наращивания (М.Н. Городний, 1980). В наших исследованиях лучшей ассимилирующей поверхностью гречихи был показатель на уровне 56,7 тыс. м2/га. Расчёты говорят о том, что в процессе фотосинтеза гречиха может формировать за сутки до 7,34 г сухой биомассы на 1 м2 листовой поверхности. При этом лучшие показатели урожайности отмечались при среднесуточной продуктивности фотосинтеза – 5,83 г сухой биомассы на 1 м2.
Таким образом, сравнение показателей ассимиляционной поверхности и фотосинтетической деятельности растений гречихи между вариантами, позволяет сделать вывод о том, что способы посева и нормы высева оказывают определяющее влияние на урожай зерна. Эти агротехнические приёмы можно отнести к наиболее результативным.
Фотосинтетический потенциал посевов гречихи является обобщающим показателем воздействия многих агротехнических факторов, в том числе способа посева и нормы высева. Н.Д. Кумскова (2004) считает, что фотосинтетический потенциал гречихи может составлять от 1,076 до 1,439 млн. м2 дней /га в зависимости от вносимых удобрений.
По мнению Н.М. Городнего (1980), основная причина отмирания большей части репродуктивных органов гречихи заключается в слабом снабжении их пластическими веществами. Невысокая озернённость гречихи обусловлена неудовлетворительным питанием её цветков из-за низкой облиственности растения и одновременного развития вегетативных и генеративных частей. Продукты синтеза перемещаются в растениях к тем органам, где они потребляются, и в том количестве, в котором они используются. Чем активнее жизнедеятельность организма и чем он мощнее, тем большее количество продуктов ассимиляции поступает в растение. Если образуется много растущих органов и общая потребность их в органическом веществе превысит синтетическую деятельность ассимиляционного аппарата растения, у такого растения наблюдается недостаток продуктов ассимиляции и его растущие части голодают. При этом больше страдают органы, которые растут менее интенсивно, в которые органические вещества поступают слабее. Их рост приостанавливается, но если недостаток питания длителен или даже постоянен, растения гибнут.
Бороздковые посевы гречихи в условиях Новосибирской области, в сравнении со сплошными рядовыми, формировали более значительный фотосинтетический потенциал. При этом преимущество бороздковых посевов по данному показателю последовательно нарастало от ранних периодов роста и развития растений, к более поздним. В среднем за 3 года исследований А.Д. Гончарова (2008), за период всходы-бутонизация, фотосинтетический потенциал бороздковых посевов был больше соответствующего показателя сплошных рядовых на 15 %, а за период бутонизация-цветение – на 28 %, за период цветение-созревание – на 30 %.
В опытах Н.Д. Кумсковой (2004) высокая облиственность растений и урожайность гречихи отмечены на посевах нормой 3 млн. всх. зёрен на 1 га. При высеве 2 млн. семян снижался выход зерна из-за недостаточной густоты стояния растений, что приводило к неэффективному использованию пашни. Нормы высева 4 и 5 млн. зёрен нерациональны, так как в таких посевах формировалась избыточная вегетативная масса. Наиболее высокая масса листьев гречихи, по отношению к массе растений (облиственность), наблюдалась в фазу бутонизации. Затем облиственность растений гречихи к началу цветения снижалась в два раза, достигала минимального значения в фазу созревания. На делянках, удобренных азотом, снижение её было более интенсивным, чем на контроле и других фонах. В этом варианте облиственность к моменту созревания равнялась 13,3 %, вместо 18,6 % на контроле.
Полученные нами данные для лесостепной зоны Алтайского края не противоречат результатам исследований Н.Д. Кумсковой (2004).
Наши наблюдения говорят о том, что максимального значения фотосинтетический потенциал достигал в разреженном травостое в фазу плодообразования, то есть при широкорядном способе посева (табл. 59).
Таблица 59
Фотосинтетический потенциал и облиственность гречихи посевной
(средние за 2010–2011 гг.)
|
Вариант |
Норма высева, млн. всх. зёрен на 1 га |
|||||
|
2,5 |
3,5 |
4,5 |
||||
|
фотосинт. потенциал |
облиственность |
фотосинт. потенциал |
облиственность |
фотосинт. потенциал |
облиственность |
|
|
Рядовой (контроль) |
1,20 |
10,1 |
1,27 |
14,4 |
1,45 |
16,0 |
|
Широкорядный (0,45 м) |
1,43 |
11,6 |
1,67 |
15,7 |
1,79 |
17,2 |
|
Широкорядный (0,60 м) |
1,70 |
13,0 |
1, 86 |
17,3 |
1,87 |
19,8 |
Примечание. Фотосинтетический потенциал – в млн. м2/сут. на 1 га; облиственность – в %.
Лучшего значения с учётом урожайности фотосинтетический потенциал достиг на междурядьях 0,45 м при норме высева 3,5 млн. всх. зёрен на 1 га и составил 1,67 млн. м2/сут. на 1 га.
Более высокая масса листьев по отношению к массе растений гречихи, в зависимости от способа посева и нормы высева, отмечена нами в фазу бутонизации – 39,7–51,2 %.
Во время цветения облиственность снижалась примерно в 2 раза на всех вариантах и минимального значения достигала в период плодообразования-созревания. Её показатели в этот период не превышали 20 %.
Таким образом, урожай гречихи определяется благоприятным сочетанием метеорологических приёмов и агротехнических условий, наиболее подходящих биологическим потребностям этой культуры. Поэтому при правильной организации агротехнического комплекса можно получить высокие урожаи зерна вследствие хороших показателей фотосинтеза.