
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

КОМПЛЕКСНАЯ ОЦЕНКА ЛЕСОВОЗОБНОВЛЕНИЯ НА ВЫРУБКАХ И ПРОЕКТИРОВАНИЕ ЛЕСОВОССТАНОВИТЕЛЬНЫХ РАБОТ
Савченкова В А,
ВВЕДЕНИЕ
Учебное пособие содержит мероприятия по проектированию мер содействия естественному возобновлению леса, посеву, посадке леса, выращиванию посадочного материала, технической приемке и инвентаризации выполненных работ. Изложена лесокультурная оценка сплошных вырубок.
Сохранение лесной экосистемы, особенно при промышленных рубках является одной из важнейших задач лесного хозяйства. Актуальной проблемой является восстановление лесных древесных ресурсов, сокращение сроков их возобновления. При подготовке лесосек к рубке и в процессе их разработок уничтожается благонадёжный подрост, имеющийся под пологом спелых и перестойных насаждений. Пассивное формирование естественных насаждений на вырубках ведёт, как правило, к существенному затягиванию сроков лесовосстановления, смене пород.
С каждым годом увеличивается площадь лесных насаждений, которые подвергаются антропогенному воздействию. Под влиянием деятельности человека происходят сукцессионные процессы. В экономике страны продукты переработки леса используются практически во всех отраслях. В связи с этим регулярное восполнение лесных ресурсов является важной производственной задачей.
Рациональное лесопользование обеспечивается соблюдением лесоводственных требований к сплошным рубкам и использованию лесных ресурсов в целом; учётом социальных и экологических функций леса для местного населения, хозяйственная деятельность которого тесно связана с лесными угодьями и лесными ресурсами; непрерывностью лесопользования и лесовосстановления хозяйственно ценными древесными породами с учётом тенденций изменения структуры лесопотребления; соответствием технических средств и технологий лесозаготовок природным особенностям леса.
Большой интерес для народного хозяйства представляют именно южнотаёжные леса, изучение природных особенностей которых и их изменений под влиянием хозяйственной деятельности человека в целях наиболее рационального использования и восстановления лесов является важной научной задачей. В связи с ростом объемов потребления древесного сырья существенное значение приобретают вопросы сокращения сроков восстановления лесных массивов до состояния эксплуатационной годности.
Актуальность вопросов лесовозобновления в настоящее время возрастает в связи с необходимостью обеспечения сырьём постоянно или длительно действующих лесопромышленных предприятий и поддержания на необходимом уровне или сохранения экологических функций леса.
Многие десятилетия в различных регионах нашей страны велись исследования площадей требующих проведения лесовосстановительных мероприятий. В результате были изучены типы вырубок, вскрыты некоторые закономерности в ходе естественного возобновления как под пологом леса, так и на вырубках, выявлены изменения живого напочвенного покрова, происходящие после рубки в некоторых типах лесорастительных условий и т.д. Полученные результаты представили практический интерес и послужили отправным материалом для дальнейшего широкого изучения вопросов лесовозобновления в связи с рубками бореальных лесов.
Проблема экологических последствий сплошных рубок (или образования типов вырубок и возобновления леса в связи с ними) в разных географических условиях освещена недостаточно. Имеющиеся сведения по отдельным регионам России касаются большей частью воздействия машин на почвенный и растительный покровы. Производственный опыт и исследования сплошных рубок свидетельствуют о том, что в условиях высокоинтенсивного лесного хозяйства мало ценится и используется способность лесов к самовозобновлению.
Сплошные рубки вызывают, с одной стороны, усиление притока солнечной радиации из-за удаления древостоя, обусловливающее микроклиматические, почвенные и прочие экологические условия, с другой – повреждение или уничтожение (частичное или полное) лесозаготовительной техникой отдельных компонентов лесного биогеоценоза (подроста, подлеска, живого напочвенного покрова, почвы и т.д.). Поэтому влияние сплошных рубок на лесные
биогеоценозы в момент их проведения слагается из воздействия техники и влияния изменённых условий места произрастания. Для того чтобы предусмотреть тенденции изменений, связанных с применением лесозаготовительной техники, необходимо их выявить и адекватно описать. Это позволит своевременно устранить или смягчить отрицательные и использовать положительные явления, вызванные воздействием сплошных рубок на экологические условия.
Промышленные рубки на территории Восточной Сибири начаты в 1949 году. До 1953 года валка леса производилась ручными пилами, а трелевка была конной. С 1953 года леспромхозы перешли на валку электропилами и трелевку тракторами. Рубки проводились концентрированные, сплошные, реже условно-сплошные (в основном до 1953 года), с непосредственным ежегодным примыканием. До 1953–1954 годы рубки по существу были бессистемные, процесс лесозаготовок был организован крайне плохо. Дровяная древесина оставлялась на вырубках в сырорастущем или срубленном состоянии, а деловая древесина вывозилась также не полностью; мелкие сортименты не имели потребления и не вывозились, очень крупные деловые сортименты не могли быть вывезены конным транспортом и также оставлялись [36, 37, 38, 39].
Действовавшие с 1968 года Правила рубок главного пользования и лесовосстановительных рубок в лесах Восточной Сибири допускали в лесосырьевых базах лесозаготовителей, закрепленных в лесах 3-й группы, лесосеки шириной 1000 м, площадью до 200 га со сроками примыкания лесосек 1–3 года. На концентрированных лесосеках резко изменяется лесорастительная среда. Как правило, обсеменение вырубок происходит на сравнительно узкой полосе, расположенной вдоль стен леса. Концентрированные лесосеки возобновились, главным образом, лиственными породами. На супесчаных почвах при наличии источников обсеменения сравнительно успешно возобновлялась сосна. При других условиях роста возобновление лесосек хвойными породами незначительное и представлено сохранившимся после рубки подростом, а также медленно появляющимся под пологом лиственных пород хвойным самосевом. При этом способе рубок наблюдалось сплошное уничтожение лесов на обширных территориях по мере строительства магистральных транспортных путей вглубь таежных территорий, особенно при машинной заготовке леса [36–39, 254–258].
Начиная с 1994 по 2007 года в регионе действовали новые, более щадящие Правила рубок главного пользования. Максимальная ширина лесосеки в 3 группе лесов принята не более 500 м, а площадь ее – 50 га. Увеличены сроки примыкания лесосек – не менее 4–5 лет. Рубка очередной лесосеки разрешается лишь при условии, что ранее вырубленная лесосека возобновилась хозяйственно ценными породами или на ней проведено искусственное лесовосстановление.
Значительная часть вырубок возобновляется естественным путем, то есть самосевом или вегетативно. Естественное возобновление на вырубках зависит не только от условий местопроизрастания, способа рубки, ширина лесосеки, но также от соблюдения технологии рубок и применяемой техники при лесозаготовках. Этому вопросу посвящены многие исследования.
Большинство исследователей [10–12, 55, 78, 232–244] считают, что уничтожение подроста, повреждение и разрушение верхних горизонтов почвы агрегатной техникой плохо отражается на последующем возобновлении. На тяжелых (влажных глинистых и суглинистых почвах применение агрегатной техники ведет к уплотнению и, как следствие ухудшению физико-химических характеристик верхнего почвенного горизонта, поэтому должно быть ограничено.Результаты опытно-производственной разработки лесосек машинами (ЛП-19 + МЛ-56,ЛТ-154) в сосняках Иркутского Приангарья по технологии параллельных лент с трелевкой на два погрузочных пункта показали, что сохранность подроста на зимних вырубках составила 52 %, на летних – 43 % [169, 190]. По данным А.И. Исаева (1981), после разработки лесосек агрегатной техникой в Приангарье на сосновых вырубках в первые 3–4 года погибает до 40–50 % сохраненного при рубке подроста. Угнетенный, появившийся под высоко сомкнутым древесным пологом подрост не имеет серьезных перспектив для дальнейшего успешного роста, и его сохранять нецелесообразно, уничтожение такого подроста агрегатной техникой не наносит вреда лесному хозяйству [24, 25, 267, 288].
Древостой с сомкнутостью крон 0,7 и ниже обусловливают относительно оптимальные условия для дальнейшего выживания подроста, который после удаления древесного полога раз-
вивается нормально [36–39].
С.Д. Титов (1991)утверждает, что наибольшее число всходов сосны приурочено к участкам с сильной степенью минерализации.
В.А. Соколов и С.К. Фарбер (1999) предлагают отказаться от трелевки по волокам, а осуществлять ее бессистемно с целью более равномерного распределения нагрузки агрегатной техники на почву. При такой технологии неизбежно будет уничтожен подрост, но, по мнению авторов нововведения, появившийся самосев сосны в местах минерализации почвы будет лучшего качества по сравнению с предварительным возобновлением. Тем самым опровергаются выводы некоторых исследователей об успешном лесовосстановлении вырубок на основе предварительного возобновления, предлагающих сохранять подрост [254–258].
При планировании сохранения подроста необходимо учитывать его возраст и физиологическое состояние, поскольку старый подрост в возрасте 25–30 лет при резком осветлении и изменении гидротермических условий после вырубки материнского древостоя в основной массе гибнет.
Вырубки сосняков разнотравных и крупнотравных возобновляются слабо, что связано как с мощно развитым травяным покровом, так и с частой горимостью.
Особенно успешно возобновление проходит на сосновых вырубках Приангарья. Здесь возобновляются сосной удовлетворительно 59 % вырубок, недостаточно – 8 %, со сменой на лиственные – 23 % и находятся в невозобновившемся состоянии 10 %. Период возобновления в среднем составляет 3–5 лет.
Возобновление проходит лучше с достаточным количеством семенников. При наличии семенников в количестве 10–15 шт./га и при куртинном их размещении удовлетворительное возобновление хвойными породами составляет 58–78 % площади. При прочих равных условиях при ширине лесосек 200–400 м удовлетворительно возобновляется хвойными породами 69 % лесосек, при ширине 500–1500 м – 45 % [36–39].
Вырубки, образованные в результате рубки кедровников, возобновляются значительно хуже, чем после рубки сосняков. В подросте кедр вытесняется пихтой, елью и лиственными породами. Удовлетворительное возобновление хвойными породами отмечено на 46 % кедровых вырубок, лиственными – 26 %, недостаточное и отсутствует – 28 %.
Вырубки, образованные в результате рубки лиственничников возобновляются хуже сосновых: удовлетворительно возобновляются хвойными породами 48 % площади, недостаточно – 14 %, со сменой на лиственные – 26 % и возобновление отсутствует – 12 % [12, 14, 24, 37, 38, 40, 41]. В разнотравной и зеленомошной группах типов леса удовлетворительное возобновление хвойными породами составляет 44–46 %, в долгомошной – до 72 %. В составе подроста преобладают сосна, ель и лиственница. Период возобновления лиственничных вырубок – 3–5 лет.
На площади вырубок, образованных в результате рубки елово-пихтовых насаждений возобновление проходит успешно. Они возобновляются хвойными породами через два года на 49 % площади, через 3–5 лет – на 54 %, через 6–10 лет – на 60 % площади. Недостаточное возобновление по истечении 10 лет отмечено всего на 4 % площадей. В подросте преобладают ель, пихта, единично кедр
[12, 14, 24, 37, 38, 40, 41, 46, 47, 48, 49].
Вырубки, образованные в результате рубки лиственных пород возобновляются хвойными породами через 6–10 лет на 38 % площади, лиственными – 50 %. Возобновление недостаточное или отсутствующее отмечено на 12 % обследованных площадей, главным образом на свежих вырубках. Помимо самосева наблюдается вегетативное возобновление.
Наиболее распространены вырубки разнотравной и зеленомошной групп типов леса. Удовлетворительное возобновление протекает на 39–47 % площади этих вырубок, в том числе хвойными породами – на 21–32 %. Возобновление в разнотравной группе типов леса происходит преимущественно лиственными породами [12, 14, 24, 37, 38, 40, 41, 46, 47, 48, 49].
Анализ микроклимата позволяет сделать определенные выводы о связях условий местопроизрастания и растительного покрова, средообразующей роли последнего.
Многочисленные исследования [98, 109, 110, 113, 119] показали, что влажность воздуха находится в строгом соответствии с погодой отдельных лет, структурой растительного покрова и обнаруживает следующие закономерности: наиболее высокая влажность в дневные часы держится на 20 см от поверхности почвы, минимальная – на высоте 200 см; разница в отдельные ясные дни достигает 34 %, тогда как в пасмурные дни она в пределах 4–13 % .
Дневные величины относительной влажности воздуха в сосняке разнотравно-коротконожковом иногда в течение месяца держатся на уровне 35–41 %. Более влажным в жаркие дни в этом типе леса оказывается слой воздуха (0–20 см) у поверхности почвы. В облачную погоду влажность его умеренна, отсюда амплитуда сезонных изменений минимальна не только по сравнению с вышележащими горизонтами, но и в сравнении с темнохвойными лесами водораздельных местоположений.
На открытом луговом участке влажность этого слоя воздуха на 3–6 % ниже, а на высоте основной массы трав она выше на 4–7 %, чем в сосновом лесу, и только на высоте 200 см величины их выравниваются [98, 109, 110, 113, 119].
При сравнении влажности воздуха с таежными участками еще резче подчеркивается специфика микроклиматического режима сосновых травяных лесов. Как в березово-пихтовом, так и в пихтово-лиственничном лесах нет большой разницы во влажности всего приземного слоя (0–200 см) воздуха.
За шесть вегетационных периодов она не превышала 15 %. По сравнению с разнотравно-коротконожковым сосняком здесь не наблюдается резких колебаний, а минимальные величины отмечены в пределах 48–51 %, которые наблюдаются в жаркие дни первой половины июля [55, 57, 65, 98, 109, 110, 113, 119].
Наиболее жесткий режим влажности воздуха создается в сосняке толокнянково-брусничном и на вырубке. Минимальная влажность отмечена в пределах 28–29 %. Суточные и сезонные изменения более контрастны, чем во всех других сообществах. В жаркие дни наиболее сухим оказывается воздух приземного слоя. В дневные часы разность во влажности воздуха на высотах 20, 50, и 200 см составляет 26–33 % на вырубке и 24–31 % под пологом леса. Однако после выпадения осадков она выравнивается во всем 200-сантиметровом слое [55, 57, 65, 98, 109, 110, 113, 119].
От количества поступающих под полог леса солнечных лучей зависит температура почвы и воздуха, а, следовательно, и процессы жизнедеятельности растений. Сосновые леса на легких почвах начинают вегетировать в среднем на 2–5 дней раньше по сравнению с травяными сосняками на тяжелых почвах. В среднем древесный полог в травяных сосновых лесах задерживает 24–37 % солнечной радиации, травостой в период полного развития – не менее 80–88 %. Под пологом сосняка толокнянково-брусничниного прозрачность древесного яруса обусловливает малый процент (около 18 %) задержания солнечной радиации. На толокнянково-брусничной вырубке он еще ниже, ориентировочно 13 %.
Сравнивая характер температурного режима воздуха в сосняке разотравно-коротконожковом и в темнохвойных лесах, можно сделать вывод, различия в температурах воздуха значительно меньше по сравнению с сосняками на легких почвах. Приземный слой воздуха под пологом соснового леса с травяным покровом на 0,6–1 °С теплее, чем в темнохвойных насаждениях.
В пихтово-лиственничном и березово-пихтовом лесах в течение всего вегетационного периода поверхность почвы является деятельной поверхностью раздела, для которой характерны наибольшие амплитуды колебаний суточных температур [54, 110, 145, 189, 192, 252].
Температура почвы в корнеобитаемом слое оказывает большое влияние на почвенные процессы, рост растений, прорастание семян. Согласно классификации В.Н. Димо для южно-таежных дерново-подзолистых почв Приангарской провинции характерен сезоннопромерзающий тип температурного режима с преобладанием талого их состояния в годовом цикле. Среднегодовая температура
воздуха –2,4°, а почвы + 2,7° [54, 110, 145, 189, 192, 252].
Температурный режим почв пространственно неоднороден в связи с рельефом, типом почв, их влажностью и структурой, а также с растительным покровом. Наблюдения ограничены тремя месяцами вегетационного периода.
Почва в темнохвойных лесах в среднем за вегетационный период на 2,1–3 °С холоднее, чем в травяных сосняках. Если принять во внимание, что корни сосны и пихты начинают расти при температуре не ниже 5-6 °, то оказывается, что в сосновых лесах в июне весь 80-сантиметровый, а в отдельные годы и 120-сантиметровый слой почвы по температурным условиям благоприятен для активной жизнедеятельности корней сосны [54, 110, 145, 189, 192, 252].
Поскольку глубина окончания вертикальных корней ограничена изотермой в 3–4°, то в травяных сосновых лесах она располагается на глубине около 200 см.
В темнохвойных лесах в июне рост корней пихты возможен только в верхнем слое почвы (10 см), в отдельные годы до 20 см в березово-пихтовом лесу и до 10, а иногда 40 см в пихтово-лиственничном. В июле этот уровень ограничен 80–100 см. Максимальное прогревание наблюдается в августе, когда изотерма 5° опускается до 200 см.
Низкие температуры почвы в июне в темнохвойных лесах задерживают рост корней пихты, в то время как температура воздуха обеспечивает возможность роста побегов. В годы с холодной и сухой весной отрицательные температуры в сочетании с недостаточной влажностью воздуха в дневные часы приводят к усыханию отставших в росте деревьев и слаборазвитого подроста [54, 110, 145, 189, 192, 252].
Ранним оттаиванием и глубоким прогреванием отличаются песчаные почвы на вырубках и под пологом леса сосняка толокнянково-брусничного. Под пологом леса в сосняке толокнянково-брусничном в течение вегетационного периода почва в среднем на 4,3–4,5 теплее, чем в травяном сосняке. Разность с вырубкой составляет 5,3–6,7°.
Дневные температуры в верхнем горизонте почвы толокнянково-брусничного сосняка и вырубки выше 30° с максимумами до 38,1–42,5 в июле являются неблагоприятным фактором для возобновления сосны. Под воздействием дневного максимума температур выше 37° у самосева и подроста нередко происходит ожег корневой шейки, что приводит к гибели самосева. Всходы сосны более чувствительны к действию высоких температур и чаще гибнут от них, чем от низких [63, 59]. Отрицательное влияние может оказывать не только температура, но и сопутствующее иссушение почвы.
К тому же в разгар вегетационного периода неблагоприятные условия для роста сосны создаются и тогда, когда температура верхних горизонтов почвы равна температуре воздуха, что вызывает подавление физиологических и ростовых процессов у растений [97, 110, 192].
Особенности водного режима почв в значительной степени регулируют жизненные процессы в растительных сообществах.
Динамика влажности почвы специфична для каждого из сообществ и изменяется в зависимости от погодных условий. Для сосняка разнотравно-коротконожкового специфика водного режима почвы в вегетационный период проявляется в периодически действующих верхних грунтовых водах. Их наличие в почвах – обычное явление для большинства типов леса таежной зоны [110, 162, 163].
В Нижнем Приангарье они свойственны преимущественно травяным сосновым лесам плоских водоразделов с суглинистыми почвами. Отрицательное влияние верхних грунтовых вод на рост и развитие древесных пород зависит от глубины их залегания, длительности затопления и от температуры почвы в период подтопления [54, 110, 145, 189, 192, 252]. После того как верхние грунтовые воды отступают они практически не играют роли в водном балансе травяных сосновых лесов.
Во время затопления корневые системы растений испытывают неблагоприятные воздействия недостатка кислорода и накопленных в анаэробных условиях восстановленных продуктов. Температура почвы в июне на глубине расположения верхних грунтовых вод не превышает 5–6°. При такой низкой температуре не происходят активные процессы жизнедеятельности корней сосны, трав и микроорганизмов, отсюда и потребность в кислороде невелика, а следовательно, и отрицательное влияние верхних грунтовых вод сводится до минимума. К тому же сосна хорошо переносит и более продолжительное избыточное увлажнение [54, 110, 145, 189, 192, 252].
Пихта особенно требовательна к богатству почв, теплу, влаге и избегает заболачивания [54, 110, 145, 189, 192, 252].
На вырубке благодаря куртинному размещению подроста и меньшей сомкнутости крон увеличивается поступление влаги в почву, что способствует большему увлажнению ее верхних горизонтов.
Периодическое высыхание верхних слоев почвы, главным образом подстилки, угнетая рост корней, подавляя деятельность полезных микроорганизмов и грибов и ухудшая снабжение деревьев элементами питания, оказывает отрицательное влияние на рост, развитие и приживаемость всходов. Отсюда в толокнянково-брусничных и лишайниково-брусничных борах наблюдается исключительно острая конкуренция между взрослыми деревьями сосны и подростом из-за влаги и питательных веществ. Аналогичное явление наблюдается и в сосновых лесах, произрастающих на легких почвах
[29, 54, 89, 100, 103, 110, 145, 162, 177, 183, 189, 192, 235, 252].
Эти воздействия недостатка влаги в травяных типах леса отражаются в основном на продуктивности травяного покрова.
Для самосева сосны, произрастающего в травяных типах, создается особенно неблагоприятная световая обстановка в первые 6–10 лет жизни, которая является причиной подавления основных физиологических процессов и приводит к нарушению обмена веществ. После выхода подроста из-под влияния трав в условиях разреженного полога древостоя не наблюдается зависимости прироста от освещенности
[29, 54, 89, 100, 103, 110, 145, 162, 177, 183, 189, 192, 235, 252].
Рост и развитие пихты в березово-пихтовых и пихтово-лиственничных лесах в значительной степени также зависит от микроклиматических условий. Крупный и благонадежный подрост расположен только в окнах древесного полога, где выше освещенность, температура воздуха и почвы.
Анализ особенностей местообитаний и микроклиматических условий сосновых и темнохвойных лесов убеждает в том, что сосна на водоразделах с дерново-подзолистыми почвами, тяжелыми по механическому, составу, находится в условиях, близких к экологическому оптимуму, тогда как пихта имеет довольно узкий экологический ареал, далекий от оптимума.
Такие особенности микроклиматического режима, как длительное сохранение сезонной мерзлоты, умеренное испарение с поверхности почвы под пологом темнохвойных лесов, хороший дренаж, а также снижение размаха колебаний температур почвы и воздуха по сравнению с водораздельными сосновыми лесами, способствуют сохранению темнохвойных лесов на высоких водоразделах в районе исследования [29, 54, 89, 100, 103, 110, 145, 162, 177, 183, 189, 192, 235, 252].
Низкая влажность воздуха в весенний период и медленное оттаивание почвы в сочетании с относительно высокими температурами воздуха ограничивают период активного роста пихты, снижают биологическую продуктивность и размеры естественного возобновления.
Таким образом, локальные различия в микроклиматических режимах, вызванные с особенностями рельефа и структурой лесных сообществ, весьма существенны количественно. В них отражается и средообразующая роль эдификаторов древесного яруса и рельефа.
Новый тип леса на месте старого может формироваться разными путями. Поэтому при изучении типов леса [128, 129, 135, 138], их использовании в практике лесного хозяйства все настоятельнее проявляется необходимость учета происхождения типа леса, значения этапов его формирования и происшедших смен, определения тенденций дальнейшего развития, понимания того, что будет в дальнейшем при естественном ходе событий, и что может быть при различных формах вмешательства человека, в том числе в смысле возможностей превращения существующего малопродуктивного типа леса в высокопродуктивный. В решении задачи создания высокопродуктивных и наиболее ценных лесов будущего, удовлетворяющих многосторонние запросы и потребности людей, динамическая типология леса может оказать большую пользу [128, 129, 135, 138].
Отечественная типология имеет длительную историю. Одним из основоположников лесной типологии в России по праву
считается Г.Ф. Морозов. Идею типов насаждений поддерживали лесовод В.Я. Добровлянский и ботаник-географ Д.М. Коржинский. К этим именам известнейших ученых следует добавить М. Турского, А.Ф. Рудзского, С.Ф. Бергера, а также лесоустроителей И.И. Гуторовича, Н. Генко, В. Ляховича, Д.Д. Назарова, Н.А. Гракова и др., которые воплотили эту идею в реальность в практической работе. Важную роль сыграло и выступление А. Крюденера на заседании Лесного общества. Он предложил понимать под типом насаждений «сумму всех факторов, дающих нам понятие об известном лесе, то есть известное сочетание климатических условий, почвы, инсоляции, состава и характера насаждения, дающее нам указание на известные условия возобновления, а поэтому и право, и обязанность ведения известного хозяйства» [122-132, 147–155]. А. Крюденер показал важность изучения растительности нижних ярусов, ее следует обязательно учитывать и использовать при выделении типов насаждений. «Подлесная флора является для нас важной помощницей при решении всех тех вопросов, на которые мы, имея дело с лесом, наталкиваемся на каждом шагу и от верной постановки которых зависит ответ, в свою очередь, влияющий на тот или другой вывод об историческом возникновении леса, о смене одних древесных пород другими, одним словом, о рождении леса, о жизни, борьбе и смерти его» [122–132, 147–155]. Тогда же А. Крюденер высказал убеждение, что со временем типы насаждений станут служить основой «при всех лесоводственных соображениях в самом широком смысле слова, включая сюда и все работы по лесоустройству.
Если Г.Ф. Морозов в своих первых типологических работах на первое место при дифференциации лесов ставил почвенно-грунтовые условия, то одновременно существовало и другое направление, которое отдавало приоритет растительности. Его основоположником был В.Н. Сукачев. Понимая лес как сообщество, приняв морозовское представление о типах насаждений, В.Н. Сукачев стремился вложить в это понятие более широкий смысл [122–123].
Тип вырубки – основная классификационная единица лесорастительных условий в пространстве и во времени применительно к сплошным вырубкам. Тип вырубки объединяет участки сплошной рубки, однородные по комплексу лесорастительных условий, характеризующиеся определенным напочвенным покровом, микроклиматическим, почвенно-гидрологическим и микробиологическим режимами, определяющими общие тенденции изменения лесорастительных условий и лесовосстановительного процесса [122–132, 147–155].
На рис. 1 представлена схема формирования типов вырубок в зависимости от типа леса в северной подзоне тайги европейской части России.
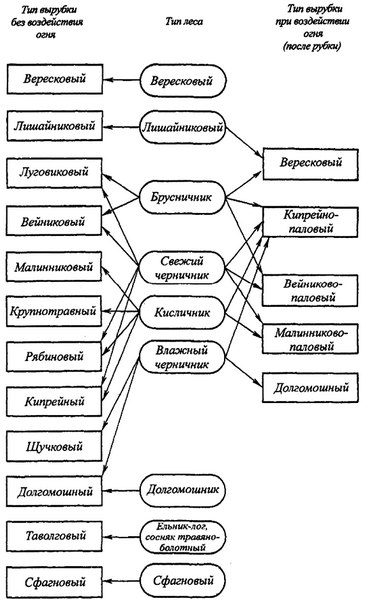
Рис. 1. Формирование типов вырубок в зависимости от типа леса в северной подзоне тайги европейской части России
Схема на рис. 1 отражает зависимость типа вырубки от типа леса и показывает:
– в одном и том же типе леса после рубки образуются разные (но строго определенные) типы вырубки в зависимости от того, подвергались или не подвергались они действию пирогенного фактора;
– типологический диапазон вырубок, образующихся на месте одного и того же типа леса, расширяется с повышением производительности (бонитета) леса.
В узких географических границах (в пределах лесничества, лесхоза) связь типов вырубки с высокопроизводительными типами леса выражена более четко [128, 129, 135, 154].
По мнению автора для разных регионов схема может уточняться и видоизменяться, не меняя своей принципиальной основы.
Облесившаяся вырубка рассматривается как начальный этап формирования типа леса. Наиболее сложившиеся черты сформировавшегося типа леса приобретаются к возрасту спелости древостоя [128, 129, 135, 154]. На сплошных вырубках лес в одних случаях формируется через смену древесных пород, в других – без смены пород. Основой для изучения типов вырубок послужила схема формирования типов леса, разработанная Мелеховым И.С. (рис. 1).
На рис. 2 можно видеть этапные смены растительного покрова вырубок, включая формирование леса через последующее
возобновление.
При установлении типа леса в натурных условиях необходимо знать происхождение типа, этапы его формирования и происшедшие смены древесной, кустарниковой и напочвенной растительности. Очень важно при этом определять тенденции последующего развития типа леса. Понимать, что будет в дальнейшем при естественном ходе событий и что может быть при различных формах вмешательства человека. Чтобы видеть динамику типов леса, вделяют отапы их развития. Они могут быть ускоренными и замедленными.
Изменения условий среды, различия в растительных этапах, их взаимосмены сказываются и на продолжительности периода возобновления леса, который в природе может быть различным. Сокращения периода для хозяйственно ценных пород – одно из важных средств повышения продуктивности лесов.
На схеме (рис. 3) показаны пути более быстрого и более медленного восстановления ельников-черничников [122–132, 147–155].
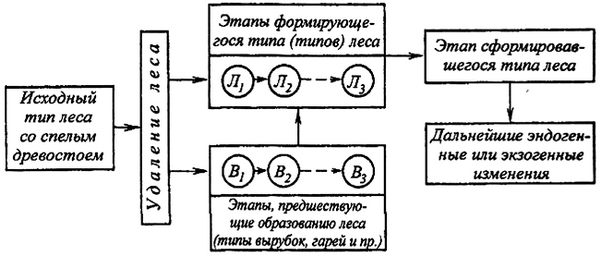
Рис. 2. Схема формирования типов леса (Л-лес, В-вырубка)
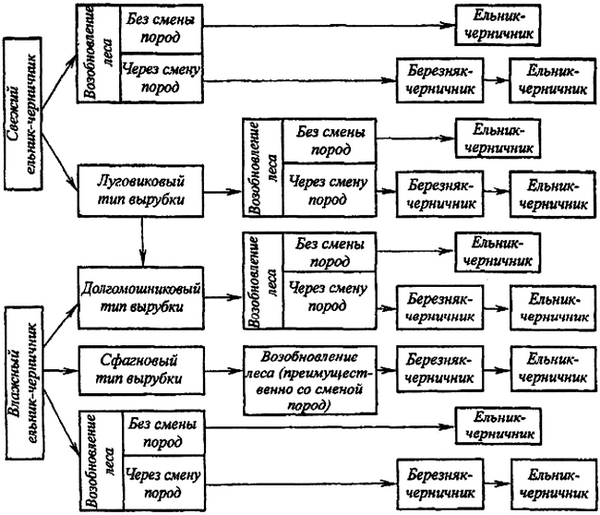
Рис. 3. Схема этапных смен растительного покрова
после рубки на месте ельников-черничников
Схема отражает возможные последствия сплошной рубки (без воздействия огня) в черничных типах леса. Соответствующие схемы могут быть составлены и для других типов леса в других географических районах. Так в некоторых районах средней тайги для сосняков – брусничников характерен этап вейникового сосняка (рис. 1).
В природе наблюдаются случаи ухудшения роста ельников при длительном существовании их на одном и том же месте. В связи с этим не исключена возможность, что в определенных случаях формирование ельника через смену березой будет более благоприятным и эффективным для продуктивности леса. Но отсюда не следует делать вывод о целесообразности повсеместной смены лиственными [133]. При этом возникает вопрос о природе новых типов леса, об их количественной и качественной продуктивности и полноценности по сравнению с лесами, поступившими в рубку. Задача состоит в том, чтобы возможно быстрее создать более полноценные древостои по сравнению с вырубленными (рис. 3).
Необходимо более определенно уточнить, в каких случаях быстрое восстановление ели без смены пород и без каких-либо промежуточных безлесных этапов не повлечет за собой снижение производительности елового леса и в каких случаях оно приведет к обратным результатам. Чтобы не допустить последних, целесообразно в определенных условиях на месте ели искусственно вводить другую хвойную породу, например, сосну (рис. 4).
Большой вклад в развитие лесной типологии внесли Е.В. Алексеев, П.С. Погребняк, Д.В. Воробьев и др. Они считали, что при выделении типов леса необходимо опираться на признаки условий местообитания. Было предложено использовать эдафическую сетку. «Типами леса, писал Е.В. Алексеев, мы называем совокупность лесных участков, сходных по климатическим и почвенно-грунтовым признакам, и, поэтому, пригодных для произрастания сходных по составу древесных пород, а часто и по покрову, основных растительных сообществ, обладающих одинаковыми лесоводственными свойствами и в силу этого допускающих применение одних и тех же мероприятий в целях возобновления и воспитания леса».
И.С. Мелехов считал важнейшей качественной особенностью типа леса его динамичность [122–132]. В его понимании тип леса – это динамическая система на биогеоценозном уровне; он характеризуется общностью морфологии, происхождения и развития лесных
сообществ, общими особенностями лесорастительных условий и тенденций дальнейшего развития леса, в процессе которого следует различать этапы, предшествующие формированию леса (типы вырубок, гарей), этапы формирующего типа леса, этап сложившегося типа леса (в спелом возрасте древостоя), возможные последующие этапы с переходом или без перехода в новый тип леса И.С. Мелехов внес большой вклад в разработку теории типов вырубок [122–132].
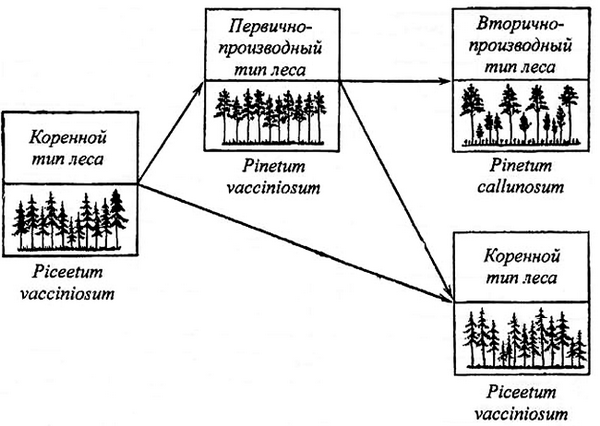
Рис. 4. Динамика ельников и сосняков-брусничников
В начале 1930-х годов, вслед за П.П. Серебрянниковым и М.Е. Ткаченко, профессор Б.А. Ивашкевич рассматривал тип леса как определенный ряд лесных сочетаний, развивающихся естественным путем в данных условиях местопроизрастания, одно из другого, в зависимости от развития древостоя, определяющего характер, этих сочетаний, то есть лишь при незначительном изменении условий местопроизрастания [129].Сопоставляя возраст модельных деревьев кедра с «пиками» числа деревьев этой породы на кривой распределения стволов по толщине, Б.А. Ивашкевич установил, что «пики» кедра обстоят друг от друга примерно на 40 лет. По его мнению, кедровый древостой состоит из ряда 35–40-летних поколений кедра, формирующихся в результате «взрывов» естественного возобновления этой породы и сменяющих друг друга в процессе развития насаждения. В онтогенезе основного лесообразователя – кедра – он выделил 8 стадий, которые последовательно проходит каждое его поколение. На разных возрастных стадиях кедр имеет различные позиции в древостое – от господства в основном пологе до участия только в подчиненных пологах [106, 129].
Лесотипологическое направление Б.А. Ивашкевича развивал Б.П. Колесников [92, 93, 129]. Он выделил классификационные единицы леса и лесорастительных условий. В динамике кедрово-широколиственного леса он выделил две фазы – преобладания в древостое кедра и доминирования его конкурентов – лиственных пород. При изучении динамики чернопихтово-широколиственных лесов Н.Г. Васильев и Б.П. Колесников [34, 106] использовали такой же подход к выделению возрастных стадий, как и в кедрово-широколиственных лесах.
Основными единицами Б.П. Колесников считает тип леса и лесную формацию. Он взял за основу схему развития кедровых лесов Б.А. Ивашкевича. Генетический подход Б.П. Колесникова к типу леса выражается главным образом в разделении древостоя на возрастные стадии. Следуя Б.А. Ивашкевичу, он уточнил схему развития кедровых лесов, приняв те же восемь стадий и 40-летнюю продолжительность каждой из их, начиная с нового поколения кедра (более 280 лет), но показав в ней одновременно стадии развития и других поколений. Таким образом, находят отражение возрастные и восстановительные стадии древостоев. Б.П.Колесников обращает внимание на необходимость изучения генезиса лесов, в первую очередь их онтогенеза, с отражением выявленных закономерностей в классификационных построениях [129].
Последующие исследователи возрастной структуры кедрово-широколиственных лесов [106] подтвердили разновозрастность кедра и образуемых им сообществах.
Н.М. Глазов [127] динамику кедрово-широколиственных лесов анализировал на уровне лесного массива Супутинского заповедника по материалам лесоустройства, полученным за 60-летний период. Он пришел к выводу, что девственный массив заповедника отличается стабильностью основных таксационных показателей (состав, запас) и развивается без смены пород.
Заключение Я.Я. Васильева [127] о вековых сменах основных лесообразующих пород на территории Уссурийского заповедника (смена кедра пихтой цельнолистной в ряде местообитаний, а также усиление позиций ели аянской и пихты белокорой в кедрово-широколиственных лесах) было основано на анализе современных фитоценотических позиций, включая естественное возобновление. Происходящие изменения в хвойно-широколиственных лесах он объяснил изменением климата, проявляющемся в увеличении влажности, в результате чего преимущество получают пихта цельнолистная и другие темнохвойные породы (ель аянская и пихта белокорая).
Интересные материалы о динамике кедрово-широколиственных лесов были получены на постоянных пробных площадях, заложенных в свое время Б.А. Ивашкевичем и его коллегами. Повторные ревизии этих площадей [122–132], а также заложенных позже, позволили отслеживать происходящие изменения в древостоях (распределение пород по толщине, состоянию и отпаду деревьев и т.д.), а также в естественном возобновлении древесных пород и делать мотивированные заключения о позициях лесообразующих видов в каждый отрезок времени. На основе этого строилось заключение о перспективах развития лесных экосистем на исследуемых участках [127].
Кудинов Ю.И., Манько И.Ю. и др., проводя исследования [113] динамики кедрово-чернопихтово-широколиственных лесов на территории Уссурийского заповедника, пришли к заключению, что на современном этапе лесообразовательного процесса наметилась четкая тенденция снижения участия Pinuskoraiensis и Abiesholophylla в сложении древостоев. Этот процесс будет продолжаться и в дальнейшем, поскольку в тонкомерной части и в подросте доля этих пород невелика и не сможет компенсировать отпад перестойных деревьев. В древостое в ближайшее время будет неуклонно возрастать роль лиственных пород, к которым перейдет преобладание в верхнем пологе. Авторы предвидят фазу господства лиственных пород, продолжительность которой трудно предсказать, так как надежного резерва для замены отмирающих поколений хвойных пород на обоих участках нет. С полной определенностью утверждают о том, что цикличность смены поколений Pinus koraiensis и Abies holophylla в изучаемых древостоях на какое-то время нарушится.
Определенный интерес представляют [120] исследования Л.Б. Махатадзе, посвященные темнохвойным лесам Кавказа (рис. 5).
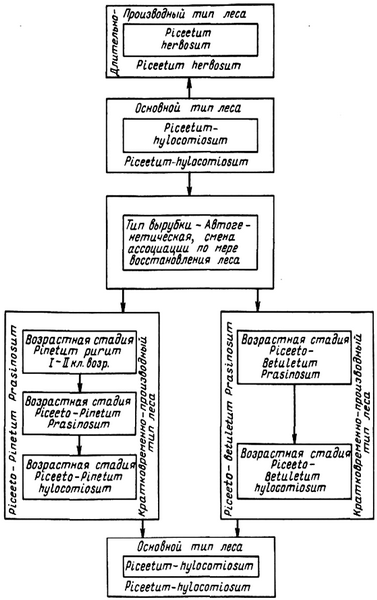
Рис. 5. Схема взаимосвязи типа условий местообитания, типов леса и возрастных стадий (ассоциаций)
Применительно к ним исследователем дана схема взаимосвязи типа условий местообитания, типов леса и возрастных стадий.
Разработана эколого-генетическая классификация типов леса.
На основе закономерной распределения типов леса в горных условиях Кавказа автор строит естественную классификацию типов леса, одним из таксонов которой является хозяйственная группа типов леса [120]. Тип леса – цикл типов леса – группа близких циклов (обычно 2–3, как исключение 1), отвечающих понятию хозсекция-биом (биогеоценотическая, или растительная, формация – дубовая, буковая, еловая и др. со всеми ее производными фитоценозами в совокупности со средой и животным миром) [120]. Биом дает возможность хозяйству установить главную целевую породу.
Разработка типологической классификации лесов, как правило, основывается на признаках вполне сформировавшихся [127], т.е. спелых насаждений. Структура фитоценоза в спелых насаждениях лучше отражает эдафические свойства местообитания, в них наблюдается наибольшее взаимосоответствие растительности и среды.
В процессе лесохозяйственной деятельности лесовод сталкивается с насаждениями различного возраста, различной стадии формирования и точное типологическое диагнозирование этих насаждений имеет не меньшее, а иногда даже большее значение, чем определение типа спелого леса [127].
Как известно, наиболее резким хозяйственным воздействием на биогеоценоз, приводящим к катастрофическим сменам, является сплошная рубка. На вырубках исчезают теневыносливые, характерные для леса индикаторные виды, формируются фитоценозы, совершенно отличные от существовавших до рубки. Правда, некоторая их связь наблюдается. Некоторые исследователи утверждают, что зачаток будущего доминирующего вида растительности вырубок имеется уже под пологом леса, где существует в угнетенном состоянии, а рубка древостоя только способствует его развитию [122–132].
Как известно, наиболее резким хозяйственным воздействием на биогеоценоз, приводящим к катастрофическим сменам, является сплошная рубка. На вырубках исчезают теневыносливые, характерные для леса индикаторные виды, формируются фитоценозы, совершенно отличные от существовавших до рубки [122–132].
Правда, некоторая их связь наблюдается. Некоторые исследователи утверждают, что зачаток будущего доминирующего вида
растительности вырубок имеется уже под пологом леса, где существует в угнетенном состоянии, а рубка древостоя только способствует его развитию [127]. Однако по мнению автора [127], для распространения того или другого вида, хотя и бывшего в зачатке под пологом, имеют значение также метеорологические условия года рубки, технология ее проведения, способ очистки лесосеки, оставляемый подрост, подлесок и другие факторы.
Автором Каразия С.П. выделено пять стадий, различающихся по густоте и составу растительного покрова, а также и по другим компонентам биогеоценоза [127], так как изменение фитоценотического состава сообществ сопровождается изменениями других компонентов биогеоценоза, а также функциональных связей между ними, что изменяет особенности процессов обмена веществ. Таким образом, следуя концепции В.Н. Сукачева, что тип леса – это тип лесного биогеоценоза, отдельные стадии развития которого можно было бы считать отдельными типами леса. Однако разделять единый лесооборазовательный процесс, охватывающий период развития одного древостоя на ряд типов леса, нецелесообразно. С позиций генетического направления лесной типологии Колесникова, а также динамической типологии Мелехова, концепции которых считаются справедливыми как с теоретической, так и с практической точки зрения, все стадии развития насаждения относятся к одному типу леса. В таком случае, считает автор, необходимо определить диагностические признаки, способствующие достоверно точно установить принадлежность отдельных биогеоценозов, отдельных участков леса, находящихся в разных стадиях возрастного развития к определенным типам леса. Не смотря на сильные изменения состава и структуры лесных сообществ в ранних стадиях возрастного развития, диагнозирование их принадлежности к определенному типу леса, отличающемуся своеобразным лесообразовательным процессом, вполне возможно, но для этого необходимы специальные исследования не только фитоценотического состава, но и других компонентов биогеоценоза, охватывающие все стадии развития насаждений [127].
Основоположник динамической типологии И.С. Мелехов подчеркнул, что эта типология отличается от других более широким подходом к природе леса, рассматривает тип леса не только в пространстве, но и во времени [122–132]. Динамическая типология включает со всей полнотой антропогенное влияние на лес,
прежде всего сплошные рубки, проводимые в настоящее время на огромных площадях.
И.С. Мелехов выделил этапы, предшествующие образованию леса (тип вырубки, гарей), этапы формирующегося типа леса (включая промежуточные, или переходные, типы леса), этап сложившегося типа леса и возможные последующие этапы с переходом и без перехода в новый тип леса [127].
В научной литературе довольно широко освещен и неоднократно дискутировался вопрос о дифференциации как составной части изреживания формирующихся древостоев. Для изучения этого вопроса многие отечественные ученые-лесоводы заложили культуры сосны различной густоты [122–123]. Определенные сводки значения густоты хвойных пород дали А.Н. Мартынов, В.В. Миронов, М.Д. Мерзленко, подчеркнув, что первоначальная густота и исходное сложение обеспечивают основные структурные элементы генезиса насаждения, определяющие ход его формирования.
Однако то, как влияет тип вырубки на дифференциацию культур, в научной литературе почти не нашло отражения. Общеизвестно, что основными причинами отпада и, как следствие, дифференциации являются наследственные свойства деревьев, заложенные в семенах, условия микросреды, куда они попали, и борьба за существование. До смыкания культур борьба внутри вида за свет, влагу и питательные вещества, можно сказать, не имеет место, а условия микросреды и борьба за существование с травяным и моховым покровом становятся одним из важнейших причин отпада и расчленения сосны по размеру в культурах [127].
Как показали исследования, выкопка и пересадка сеянцев хвойных пород вызывает торможение роста надземной части культур, которое продолжается в течение двух лет [12, 13, 15, 18, 148]. В этот промежуток времени (в первый год) происходит преимущественно регенирация корней, а затем (во второй год) общее нарастание хвои и корней как основы для увеличения прироста. Одновременно в этот период происходит формирование любого типа вырубки. На 2–3-летних вырубках травяной покров вступает в начальную фазу биологического расцвета, а культуры сосны находятся в фазе торможения роста. Поэтому, когда культуры на 3-й год после их закладки вступают в период быстрого роста, они уже находятся под заглушающим пологом травянистого покрова [148].
Облесившуюся вырубку с сомкнутым молодняком, как отмечает И.С. Мелехов, следует рассматривать как начало лесного этапа
[7–9, 122–132, 148]. Процесс возобновления, как правило, завершается образованием сомкнутого молодняка или чащи, наступает начальный этап становления леса.
В итоге при формировании типов леса после сплошной рубки сосновых древостоев и создания в первые пять лет культур сосны можно выделить следующие наиболее важные этапы [127]:
– этап, предшествующий образованию типа леса (тип вырубки);
– начальный этап формирования типа леса;
– заключительный этап образования типа леса;
– этап сложившегося типа леса;
– этап последующих эндогенных и экзогенных изменений в типе леса.
Каждому этапу [127], особенно типу вырубки и начальному этапу в образовании нового типа леса на месте старого под влиянием антропогенных факторов, присущи характерные черты и особенности, связанные с приживаемостью сосны в культурах, распределением числа деревьев по классам роста (по Крафту), динамикой живого напочвенного покрова, заселением вырубок и культур лиственными и хвойными породами, формированием состава древостоев, продуктивностью сосняков искусственного происхождения и т.д.
Мерзленко М.Д. утверждает, что искусственные насаждения представляют собой незаменимые объекты для экспериментальных исследований в области лесной экологии и динамической типологии леса [134]. Автором была рассмотрена последовательность чередований этапов (стадий), возникающих в процессе формирования, ельника-кисличника искусственного происхождения включительно по 100-летний возраст. Для культур ели было выделено пять этапов (рис. 6).
Рассматривая всю их совокупность, слагающих формирование и развитие ельника-кисличника искусственного происхождения, в свете динамической типологии установлено явление цикличности этапов в формировании и развитии данного типа леса [134]. Оно носит характер замкнутого цикла с четкой последовательностью чередования ряда определенных этапов.
Такой замкнутый цикл происходит при целенаправленной деятельности человека, сводящейся к созданию в условиях коренных ельников-кисличников культур ели.
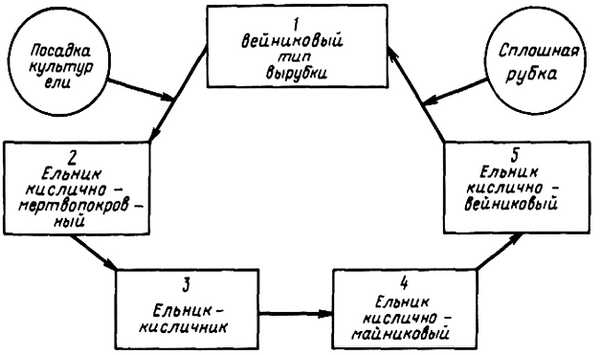
Рис. 6. Цикл этапов в формировании и развитии ельника-кисличника искусственного происхождения
Учение о динамической типологии, научные основы которого разработаны И.С. Мелеховым, позволяет наиболее правильно прогнозировать рост и развитие древостоев и на его основе решать практические задачи лесного хозяйства. Важное значение динамическая типология имеет в обширной области антропогенных смен, вызываемых многообразной деятельностью человека, особенно при разрешении вопросов, связанных с рубками леса, лесовосстановлением, продуктивностью лесов.
Учение о типах вырубок, как и учение о типах леса, возникло в России. Типология вырубок, созданная академиком И.С. Мелеховым [122–132], имеет более чем полувековую историю, которую следует рассматиривать как становление и развитие исследований природы вырубок в лесоводственно-биогеоценитическом и лесоводственно-географическом планах [118, 127, 136, 150].
И.С. Мелехов [127] вырубку (как и лес) рассматривал как природное единство (биогеоценоз), где вся растительность, фауна, микроорганизмы, почва и атмосфера находятся в тесном взаимодействии. В связи с этим в нашей стране широко осуществлялось комплексное, лесоводственно-биогеоценотическое изучение природы вырубок и возобновление леса [127, 141, 156]. Значительное внимание при этом удилялось исследованию живого напочвенного покрова, его индикаторной и идификаторной роли в различных экологических условиях, который является наиболее наглядным внешним показателем природного единства в условиях сплошных рубок [127, 141, 156]. По мнению И.С. Мелехова «в растительном покрове необходимо выделять такие явления, происходящие после рубки, которые наглядно отражая определенный этап в изменении экологических условий, имеют наибольшее значение для практики» [127]. Так, разные виды напочвенного покрова неодинаково влияют на лесоратительные условия сплошных врубок. Например, большинство видов вейника создают сильное задернение поверхности почвы после сплошных рубок, затрудняющие возобновление хвойных пород. Иван-чай узколистный, наооборот, способствует образованию благоприятных условий для лесовозобновления. Заросли последнего образуют благоприятные микроклиматические условия для появления всходов, роста самосева и подроста. Надземная часть этого вида, быстро разлагаясь, повышает плодородие почвы. Одним из важнейших индикаторов заболачивания почвы является сфагновый мох, который в тоже время служит эдификатором (созидателем) лесорастительных условий, то есть способствует прогрессивному заболачиванию почвы. Следовательно, преобладающие виды живого напочвенного покрова на площадях сплошных рубок являются одновременно индикаторами и эдификаторами лесорастительных условий на сплошных вырубках. Поэтому академик И.С. Мелехов типы вырубок назвал по преобладающему виду живого напочвенного покрова, при этом рассматривается биогоценоз, охватывающий все его компоненты (растительность, животный мир, мир микроорганизмов, почву, микроклимат), находящиеся
во взаимодействии [122–132].
Известно, что для успешного возобновлния леса необходимо две группы условий: наличие благоприятных экологических условий для появления всходов, роста и развития подроста и наличия достаточного количества высококачественных семян. Даже на вырубках типов с благоприятными лесоратительными условиями при отсутствии источников обсеменния полноценное возобновление леса врядли возможно. Поэтому на вырубках одного и того же типа вырубки могут образовываться разные типы возобновления
и типы формирования леса.
Типология вырубок является прежде всего современной научной и практической основой лесовозобновления. Это можно показать на следующем примере, проводимом И.С. Мелеховым при чтении одной из лекций для студентов по дисциплине «Лесоводство». В качестве объекта исследований служил бор (сосняк) брусничный (по В.Н. Сукачеву) или А2 (по П.С. Погребняку) в северной тайге европейской части России. Изучались лесоратительные условия и лесовозобновление на трех участках: в лесу, на вырубке и на вырубке, пройденной палом (пожаром). На всех этих участках тип леса по калссификации П.С. Погребняка был А2. В соответствии со схемой формирования типов леса в северной и средней тайге европейской части нашей страны после сплошной рубки в сосняке брусничном образуется луговиковый тип вырубки, а если после такого способа рубки проходит низовой пожар – кипрейно-паловый тип [176, 185]. В условиях луговикового типа вырубки луговик извилистый как эдификатор условий среды создает сильное задернение почвы, способствует образованию мощного слоя подстилки, тем самым формирует неблагоприятные условия для возобновления сосны. На участке вырубки кипрейно-палового типа иван-чай узколистный не создает заметного задернения почвы, опад из отмерших его частей хорошо разлагается, тем самым повышая плодородие почвы. Условия для возобновления сосны здесь создаются благоприятные. Следовательно, лесорастительные условия для возобновления сосны на всех трех участках (в лесу, на вырубке и на вырубке, пройденной низовым пожаром) неодинаковы. А по классификации А.С. Погребняка на вышеуказанных трех участках тип леса один – А2. Согласно классификации И.С. Мелехова, как было отмечено выше, формируются луговиковый и кипрейно-паловый типы вырубок с разными условиями для возобновления главной породы. Теоретическое значение типологии вырубок, как научной основы лесовосстановления, состоит в частности в том, что она позволяет (по сравнению с классификацией П.С. Погребняка) более корректно устанавливать тип лесорастительных условий на площадях сплошных рубок.
Практическое значение типологии вырубок (в этом случае состоит в том, что, зная возможные изменения экологических условий (формирование того или иного типа вырубки в связи с исходным типом леса) и этапы возобновления леса, можно начинать оптимальные или хозяйственно-целесообразные решения
по лесовосстановлению. В частности, (применительно к приведенному примеру) на месте сосняка брусничника после рубки необходимо планировать лесные культуры, а после пожара на вырубках (того же исходного типа леса) ориентироваться на естественное возобновление главной породы.
Типология вырубок представляет научный и практический интерес и при разработке лесоводственно-экологических требований к работе лесозаготовительных машин на лесосеках без подроста. Методический подходк обоснованию критериев такой оценки работы техники при сплошных рубках основан на использовании показателей состояния лесной экосистемы с учетом мозаичности растительного покрова. Особое значение для установления критериев имеют входные показатели (степень минерализаии почвы, плотность ее верхних слоев), существенно влияющие на экосистемы – типы вырубок, начальный и последующие этапы формирования типов леса.
При проведении сплошных рубок на лесосеках без подроста, отнесенных к участкам с последующим способом возобновления после рубки, следует допускать образование фрагментов типов вырубок (с благоприятными условиями для возобновления главных пород в формирующихся в них парцеллах растительного сообщества) таких размеров и встречемости на площадях, которые позволили бы обеспечить встречаемость самосева и подроста для полноценного и своевременного восстановления леса на вырубке [27, 28]. А.И. Бузыкин и Л.С. Пшеничникова [184] аналогичные парцеллы относят к микросистемам с «откликающимися» хорошей возобновляемостью и фактом поселившегося самосева.
Формолизовать лесоводственно-экологические требования к работе лесозаготовительной техники на лесосеках без подроста труднее, чем в насаждениях с подростом [37, 40, 43, 48, 52–58, 60, 62, 77, 79, 93, 110–111, 127]. Сложность решения этой задачи состоит в том, что мы имеем дело не с реальным (до рубки), а с будущим (последующим) возобновлением леса, которое сразу же после рубки прогнозируется с определенной достоверностью исходя из состояния лесорастительных условий на фрагментах свежих вырубок и формирующихся на них парцеллах растительных сообществ при наличии источников семян. Поэтому для объективной оценки работы лесозаготовительной техники необходимы научные сведения для разных эколого-географических условий о характере повреждения
почвенного покрова в связи с применением того или иного типа машин и технологий, о природе и развитии парцелл и типов вырубок, об их влиянии на появление всходов и формиирование самосева и подроста. Данные сведения по ряду регионов имеются.
В практике лесоводственно-экологической экспертизы работы лесозаготовительных машин иногда могут возникнуть затруднения в определении фрагментов вырубок и образованных на них парцелл с разным состояниемэкологических условий, вызванных, например, отсутствием приборов для установления плотности почвы или непосредственной информации о возможной стпени задернния ее или другими причинами. В таких случаях для определения экологического состояния фрагментов вырубок или парцелл в ачестве придержек могут быть использованы экспериментальные или научные сведения из литературных и других источников, касающихся элементов лесосек и вырубок с благоприятными и неблагоприятными условиями для возобновления леса. Например, в ельнике черничном свежем и близком к нему ельнике кисличном зоны смешанных лесов Русской равнины после сплошной рубки всю поверхность вырубки следует отнести к неблагоприятной для возобновления главной породы из-за сильного задернения ситником и вейником как на поврежденной, так и на неповрежденной почве [152, 156, 159]. В сосняке брусничном и близком к нему по экологическим условиям типах леса Средней и Южной тайги Сибири поврежденная поверхность почвы (за исключением магистральных волоков, погрузочных площадок и лесовозных усов), в связи с незначительным задерннеим, сравнительно благоприятны для возобновления главной породы [159].
При определении размера главного пользования (для проведения рубок в спелых и перестойных лесных насаждений – согласно Лесному Кодексу РФ) не учитывается безлесный этап (тип вырубок) после сплошной рубки. На это впервые обратил
внимание профессор А.В. Вагин.
Без учета безлесного этапа после рубки невозможно с достаточной определенностью прогнозировать динамику леса и, следовательно, – объективно определить размеры главного пользования лесом. Типология вырубок позволяет решить такую сложную проблему. Для этого необходимо использовать региональные схемы формирования типов вырубок, разработанные на научно-методическом подходе академика И.С. Мелехова [127].
Наиболее эффективны мероприятия по сохранению и восстановлению дикоратущих ягодников (брусники, черники) после сплошных рубок в тех случаях, когда они планируются и осуществляются не только с учетом типов леса, но и формирующихся типов вырубок.
При оценке пожарной опасности на вырубках используется критеий горимости всего диапазона типов вырубок определенных природных зон. В последнее время теоретическая и практическая значимость типологии вырубок возрастает в связи с задачами возобновления леса на гарях, механизации лесокультурных работ, выбора лесозаготовительной техники и технологии лесосечных работ, таксации вырубок, сельскохозяйственного их освоения и прочих факторов.
Для теории и практики лесного хозяйства имеют важное значение географические аспекты типологии вырубок. К настоящему времени разработан ряд региональных схем формирования типов вырубок в связи с исходными типами леса (для европейской части России [136, 150, 291], Урала [76, 90, 99, 156], Западной [103, 159] и Восточной Сибири [141, 157, 229] и для Дальнего Востока [118, 158, 160]. Такие схемы являются современной научной и практической основой лесовосстановления. Значительно возросла изученность типов вырубок, в частности, образованных после работы агрегатной лесозаготовительной техники [156, 157]. Для отдельных регионов установлена причинно-следственная связь между степенью воздействия лесозаготовительной техники на компоненты лесного биогеоценоза (подрост, почву и др.) и формирующимися типами вырубок.
Типология вырубок позволяет при проведении лесовосстановительных мероприятий своевременно использовать положительные и свести к минимуму негативные последствия рубок. Следовательно, учение о типах вырубок И.С. Мелехова направлено на поддержание и повышение устойчивости и повышение устойчивости лесов, лесов, которую Г.Ф. Морозов [122–132] считал «верховным» принципом лесоводства. И в свою очередь это учение является лесоводственной основой устойчивого ведения лесного хозяйства.
Проектируемые способы работ и принимаемые технологические решения по лесовосстановлению должны обеспечивать своевременное воспроизводство лесов, улучшение породного состава, увеличение производительности лесов, рациональное использование земель лесного фонда. Установление объемов лесовосстановительных работ, способов их проведения и определение воспроизводимых древесных пород осуществляют лесничества в соответствии с лесоустроительными проектами с учетом изменений, происшедших и лесном фонде. Работы по лесовосстановлению и выращиванию посадочного материала осуществляют лесхозяйственные организации областного органа управления лесным хозяйством и лесопользователи. В связи с этим содержание учебного пособия изложено в стиле, максимально приближенном к действующим законодательным актам, и направлено на подготовку студентов к работе на производстве.
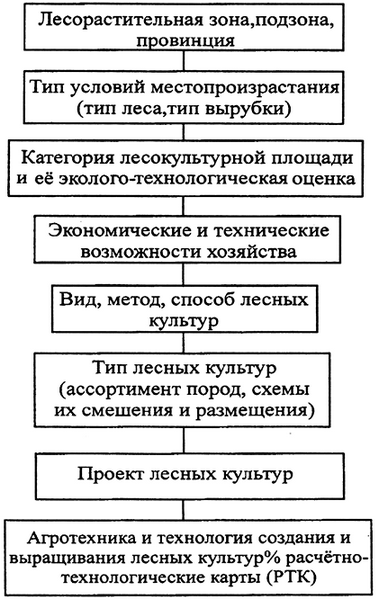
Рис. 7. Последовательность рассмотрения факторов, определяющих проектирование лесных культур
Важнейшим стратегическим направлением при выращивании искусственных насаждений на каждом участке является экологическое обоснование выбора агротехники, технологии и типа лесных культур – экспертиза на экологическую чистоту проекта [188]. Основой фундамента искусственного лесовосстановления и лесоразведения по мнению профессора Родина А.Р. являются лесорастительное районирование и лесная типология. В связи с этим планирование и проектирование лесных культур начинается с изучения лесорастительной зоны соответствующего региона (рис. 7). Это позволяет установить почвенно-климатические условия региона, произрастающие здесь древесные породы и другую растительность, а также выявить факторы (положительные и отрицательные – весенние заморозки и т.п.), влияющие на лесные древесные породы. Этапы, отраженные на схеме (рис. 7) рассматриваются в настоящем учебном пособии.
В соответствии со статьей 61 ЛК РФ вырубленные, погибшие, поврежденные леса подлежат воспроизводству. Воспроизводство лесов осуществляется путем лесовосстановления и ухода за лесами.
Лесовосстановление обеспечивается:
1. На лесных участках, предоставленных в аренду для заготовки древесины арендаторами этих лесных участков;
2. На лесных участках, за исключением указанных в подпункте «а», органами государственной власти, органами местного самоуправления в пределах их полномочий, определенных в соответствии со статьями 81–84 ЛК РФ.