
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

7.2. Сбег ствола материнского дерева и рост потомства
В насаждениях складывается разная история их развития. Контрасты в начальной густоте вполне могут привести к формированию ценозов, отличающихся генетически, и их генофонд будет передан следующему поколению. Исследований по этому вопросу у лесных генетиков крайне мало. Пока известны лишь работы А. М. Голикова (2011, 2014).
Воздействие конкуренции в прошлом отражает, а если выразиться образно, регистрирует и «записывает на себя» сбег ствола. Чем выше начальная густота, тем меньше сбег стволов; он тонко реагирует на нее и является индикатором, по которому можно реконструировать историю густоты. Например, зная таксационные показатели и густоту древостоя сейчас и 20 лет назад, можно найти такой же сбег ствола у других древостоев и использовать его для определения их густоты в прошлом. По этому принципу можно восстановить историю развития древостоя с определением его плотности в ранних возрастах и «добраться» до ее истоков даже у спелого древостоя. Именно так подбирались «естественные ряды» при составлении таблиц хода роста с разной начальной густотой по оригинальной методике Г. С. Разина (Рогозин, Разин, 2015). Сбег ствола чрезвычайно важен для характеристики общего состояния древостоя, и в формуле оптимальной густоты выращивания древостоев (Разин, 1989, а.с. на изобретение SU 1464970) сбег ствола использован дважды.
Сбег ствола у плюс-деревьев в естественных популяциях составил в среднем 1,43 см/м против 1,20 см/м в культурах. Различия вызваны как более старшим возрастом деревьев в естественных лесах, который несколько увеличивает сбег, так и большей густотой культур, снижающей его. Между 12 ценопопуляциями средний сбег ствола различался недостоверно, так как объемы выборок оказались недостаточны. Поэтому плюс-деревья объединили в 2 группы: группу из естественных ценозов и группу из культур. Влияние сбега ствола на потомство показано на двух полях корреляций (рис. 7,6 и 7,7).
Результаты весьма интересны. У плюс-деревьев из естественных ценозов в градации среднего сбега (1,22–1,51 см/м) частота лучших семей повышена в 1,9–2,4 раза в сравнению с деревьями с малым (1,0–1,21) и большим (1,52–1,9 см/м) сбегом, при некотором слабом снижении высоты семей от малого сбега к большому, судя по линии тренда (см. рис. 7.6).
У плюс-деревьев из культур зависимость аналогичная. У матерей со средним сбегом, который в этих, более густых ценозах, значительно меньше (1,1–1,3 см/м) точно также отмечено повышение частоты лучших семей в 1,5–1,9 раза по сравнению с деревьями с большими или меньшими значениями сбега. Линия тренда здесь показывает практическое отсутствие какого-либо влияния сбега на высоту семей.
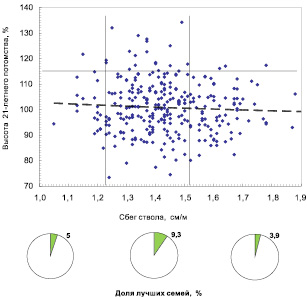
Рис. 7.6. Разделение деревьев из естественных популяций по сбегу ствола на три градации и доля в них лучших семей (с высотами от 115 %)
Для этих двух совокупностей, однако, имеется и совпадение параметров сбега, в которых наблюдается наилучшее качество потомства: это значения 1,2–1,3 см/м. Данный параметр можно рекомендовать как общую придержку для отбора плюсовых деревьев ели финской во всех популяциях.
Эффект этой простой рекомендации весьма значителен: в потомствах плюс-деревьев со средним сбегом ствола, имеющего четкие параметры, частота лучших семей повышена в среднем в 1,9 раза.
Сбег ствола отражает коллизии конкуренции (в главе 3 мы говорили о том, что он «записывает» историю конкуренции дерева) и характеризует, вероятно, генетические особенности и ценоза, и отдельных деревьев. В условиях высокой густоты и при наблюдаемых малых значениях сбега ствола, например, 1,0–1,2 см/м, в древостое накапливаются родители, толерантные к конкуренции и его генофонд можно назвать «конкурентным».
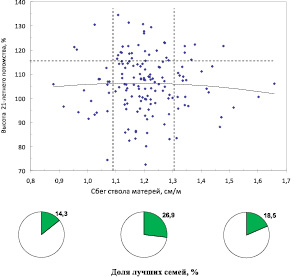
Рис. 7.7. Разделение деревьев из культур по сбегу ствола на три градации и доля лучших семей (с высотами от 115 %) в этих градациях сбега