
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

7.11. Селекция по хемомаркерам
Известны работы, где найдены существенные различия в содержании химических элементов в хвое у клонов сосны обыкновенной, и они коррелятивно связаны со скоростью роста и генетическими отличиями (Тараканов и др., 2007). Детально этот вопрос нами изложен в монографии (https://elis.psu.ru/node/440267), где показаны корреляции состава почвы вблизи каждого из 107 изученных деревьев с химическим составом их хвои. Кратко поясним, что почва в этих 107 образцах оказалась однородной, и содержание химических элементов в ней с точно теми же элементами в хвое достоверно не коррелировало. Приведем отдельные моменты этого исследования.
Методика отбора образцов хвои на химический анализ состояла в следующем. На делянке выбирали одно типичное дерево от семьи, и в средней части кроны нарезали около 300 г ветвей с 1–3-летней хвоей. Образец хвои брали только на участке с редкими культурами. Далее хвою сжигали в муфельной печи и проводили химический анализ золы на масс-спектрометре с индуктивно-связанной плазмой «ELAN 9000» в Институте промышленной экологии Уральского отделения РАН, с определением 32 элементов.
Анализ полученных данных показал следующее.
Зольность хвои в потомстве культур, называемых здесь сокращенно «потомство К», составила 5,31 ± 0,11 г/кг, а в потомстве естественных популяций, называемых «потомство Е» – 6,03 ± 0,10 г/кг. Увеличение составило 0,72 г/кг или 13,6 % при t = 4,9 > t0,05 = 1,98. При этом вариация зольности хвои составила для потомств Е и К, соответственно, 12,4 и 14,5 %.
Напомним, что материнские насаждения имели существенные различия по размерам шишек: культуры тяготели к ели европейской, а естественные популяции – к ели сибирской. Повышенная зольность хвои у последних популяций полностью согласуется со сведениями о том, что фенотипы с преобладанием признаков ели сибирской имеет более плотную древесину (Коренев, 2008). Вероятно, это должно обуславливать лучшую устойчивость более «сибирских» популяций ели финской к резко континентальному климату, т.е. к сильным морозам.
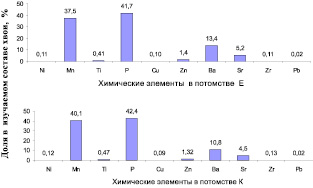
Общая картина соотношения концентраций химических элементов (ХЭ) в хвое оказалась одинакова в потомствах Е и К, с полным совпадением рангов наиболее весомых из них. Наибольший вклад вносят фосфор (P) и марганец (Mn), далее идут Ba, Sr и Zn (рис. 7.18).
Для представления о различиях в накоплении ХЭ в хвое потомств Е и K использовано вначале простое их сравнение, а именно, разность средних значений ХЭ в потомствах Е и K, принимая за 100 % их содержание в потомстве Е: (Е – K)/Е×100. В результате оказалось, что в потомстве естественных популяций содержание семи элементов оказалось выше, а трех – ниже, чем в потомстве культур (рис. 7.19).
Далее необходимо было определить достоверность превышений, и здесь возникли сложности, связанные с правомерностью использования стандартных статистических процедур. Обычно определяют стандартное отклонение, рассчитывают ошибку среднего, определяют критерий различия между средними (t) и сравнивают его с принятым уровнем безошибочного прогноза (t0,99; t0,95 или t0,90). Однако сама эта процедура допускается в строго определенном случае, а именно, при наличии двух условий.
б
Рис. 7.18. Содержание химических элементов в хвое потомства естественных популяций (а) и в потомстве лесных культур (б) ели финской

Рис. 7.19. Превышения содержания химических элементов в хвое потомства из естественных популяций в сравнении с потомством из культур
Первое условие состоит в том, что дисперсии в выборках должны быть равны и отсутствие различий между ними доказывают с помощью критерия Фишера F по отношению дисперсий (Айвазян, Мхитарян, 1998).
Второе условие заключается в распределении частот, достоверно не отличающемся от нормального. Если распределение иное и имеет, например, сильную асимметрию, то требуется преобразование данных в иную шкалу (например, логарифмическую). Отличие от нормального распределения оценивают с помощью критерия «хи-квадрат» (χ2).
Анализ данных о росте 107 семей и химическом составе их хвои включал анализ 15 полей корреляции, 22 рядов распределения частот элементов, а также данных из 13 больших таблиц, включая 5 приложений в упомянутой монографии (https://elis.psu.ru/node/440267, и база данных состояла из более чем 3 тыс. измерений. По сути, селекция по хемомаркерам становится специальным разделом селекции и требует отдельного изучения. Поэтому укажем только ее главные результаты, полученные в Пермских популяциях ели.
Так, для примера рассмотрим влияние суммы ХЭ в хвое на рост семей в редких и густых тест-культурах, поместив поля корреляции рядом (рис. 7.20).
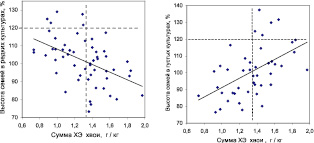
Рис. 7.20. Влияние суммы концентраций химических элементов в хвое
на рост семей ели в редких (слева) и в густых тест-культурах (справа)
На участке редких культур слева, где брали образцы хвои, связь между суммой ХЭ и ростом семей отрицательна (r = –0,47 ± 0,10), тогда как в густых культурах на правом поле связь стала положительной (r = 0,51 ± 0,11). Но на правом поле хвою на анализ не брали, и поэтому объяснить здесь влияние суммы ХЭ на рост семей можно, предполагая, что этот ее химический состав определяется генотипом.
Последнее является гипотезой, но эта гипотеза основана на экспериментальном факте – двух диаграммах (см. рис. 7.20), где зафиксированы противоположные связи. Конечно, если мы будем стоять на доктрине накопления ХЭ в тканях растения в зависимости от их содержания в почве, то тогда график справа нам покажется не имеющим смысла, так как он находится на участке, где хвою не брали на анализ. Но А. Л. Ковалевский (Ковалевский, 1983, 2003) показал, что генеративные органы, листья и побеги растений являются не-концентрирующими ХЭ частями растения (концентрируют их корни и опробковевшие ткани, например, кора). Поэтому в нашем случае хвоя – не-концентрирующий ХЭ орган растения. Более того, для нее было доказано, что изменчивость ХЭ в хвое варьирует самостоятельно и практически не зависит от их содержания в однородной по составу почве (Рогозин, Жекина, 2016; Рогозин и др., 2017; https://elis.psu.ru/node/440267). Поэтому гипотеза о генетической обусловленности химического состава хвои имеет под собой достаточно оснований, и можно судить о влиянии этого признака (хемопризнака) на рост потомства точно также, как о влиянии, например, массы семени или других морфометрических признаков потомства на его рост.
Обладая разным химическим составом хвои, семьи имеют, безусловно, разную физиологию и, как это следует из рисунка, совершенно по-разному растут в редких и в густых культурах. В редких культурах более успешны семьи с меньшим, а в густых – с большим содержанием ХЭ в хвое. Поэтому можно полагать, что разное содержание химических элементов в хвое проявляют себя как хемопризнак, имеющий наследственную основу.
Для оценки эффекта использования ХЭ в селекции можно имитировать отбор семей по сумме ХЭ (имитация отбора – вертикальная линия на графиках), ориентируясь на значение 1,35 г/кг, и отбирать в редких культурах семьи с малым, а в густых – с большим содержанием суммы ХЭ. После этого частота быстрорастущих семей (выше горизонтальной линии) составит на левом поле 5/27 = 0,185,
и по отношению к исходной их частоте, равной 5/55 = 0,091, новая частота окажется выше в 2,03 раза. На правом графике исходная их частота была 5/44 = 0,114, а новая оказалась 5/24 = 0,208, что выше исходной в 1,82 раза. Отобранные по указанным критериям семьи имеют среднюю высоту в редких культурах 108,4 ± 2,2 %, а в густых 107,2 ± 3,1 %. Это неплохой результат для коррелятивной селекции. И еще. Если учесть, что для получения образцов хвои растения можно выращивать всего лишь 3–4 года, то в том случае, если на сумму ХЭ не будет влиять возраст растений, что следует проверить в дополнительных опытах, желательные семьи можно будет выявлять очень рано.
Следует особо отметить, что сумма ХЭ влияет на рост потомства намного сильнее, чем высота, диаметр, масса семени и другие показатели материнского дерева, имитация отбора по которым повышала высоту потомства не более чем на 3,0 %, о чем уже шла речь выше. Селекция с использованием хемопризнаков только началась, но перспективы обнадеживают. Так, сумма 12 изученных элементов тесно связана с зольностью хвои (r = 0,72 ± 0,07), которую определяют довольно просто при подготовке пробы к анализу, и она влияет на рост семей примерно так же, как и сумма ХЭ на отмеченном выше рисунке. Соблюдая определенные процедуры, которые являются «ноу-хау» автора, и при использовании определенных параметров зольности хвои частоту лучших семей можно повысить в 3,4 раза (Способ повышения встречаемости быстрорастущих семей…, 2016).
В развитие идеи отбора по хемопризнакам отметим, что химический «портрет» лучших семей имеет некие особые концентрации ряда элементов, свойственных почти исключительно им. Их набор совершенно разный – при выращивании в редких тест-культурах он один, а при выращивании в густых культурах он совершенно другой, и этот набор элементов является предметом патентования. Обнаруженные различия в этом наборе элементов логически вытекают из разной картины рассеивания точек на правом и левом полях (см. рис. 7.20). С теоретических позиций эти различия вполне объясняет эпигенетика, когда условия среды влияют на экспрессию генов и далее на фенотип и его физиологию. В целом это проявление взаимодействия генотип-среда и влияния на фенотип лимитирующих факторов среды (Драгавцев, 2013), из которых, как было показано выше, наиболее сильна густота. Эпигенетические влияния были известны давно, но только в последнее время появились методы их изучения их на уровне генома (Крутовский, 2015).
Далее, и это кажется невероятным, но в потомстве, полученном от плюс-деревьев из популяций-аналогов плантаций (из культур Теплоухова), имитация отбора по таким отдельным химическим элементам позволила отобрать группу из 6 семей со средней высотой 133 % от контроля (!). При этом частота лучших семей в этой группе составила 83 % (из 6 семей 5 семей оказались с высотой от 115 %). То есть некоторые химические элементы при вполне определенной их концентрации в хвое выступают уже как хемомаркеры, и по ним можно опознавать семьи с быстрым ростом с вероятностью до 83 %. В потомстве естественных популяций такая же процедура отбора по хемомаркерам, описанная в монографии (https://elis.psu.ru/node/440267) позволила опознать семьи с быстрым ростом с вероятностью 50 %. Напомним, что исходная вероятность выделения быстрорастущих семей, обычно называемая в лесной селекции «интенсивность отбора», составляла в этом опыте 10 %.
Все эти новейшие сведения говорят о том, что ель отнюдь не испытывает дефицита по изученным химическим элементам в типичных для нее условиях в противовес расхожему мнению, что растениям может «не хватать» каких-то элементов. Наоборот, их малое содержание в хвое, обусловленное генетически, повышают продуктивность особей в культурах с малой густотой, что как раз и нужно для выращивания ели на плантациях с укороченным оборотом рубки.
Выводы
1. Семьи, хорошо растущие в редких культурах, содержат в хвое суммарно в 1,28 раза меньше Ni, Cr, Mn, V, Ti, P, Cu, Zn, Ba, Sr, Zr, Pb, чем семьи, успешно растущие в густых культурах.
2. Влияние суммы химических элементов в хвое на рост семей в редких и густых культурах совершенно разное. В редких культурах, где хвою брали на анализ, связь между суммой химических элементов в хвое и ростом семей отрицательна (r = –0,47 ± 0,10), но в густых культурах (где хвою на анализ не брали) влияние положительное (r = 0,51 ± 0,11). Поэтому можно полагать, что химический состав хвои мало связан с условиями среды, обусловлен генетически и является для семьи ее хемопризнаком.
3. Имитация отбора семей с суммой содержания химических элементов в хвое менее 1,35 г/кг увеличивала их высоты в редких культурах до 108,4 ± 2,2 %. Напротив, при отборе по их сумме более 1,35 г/кг семьи увеличивали высоту только в густых культурах (до 107,2 ± 3,1 %). Частота лучших семей при такой имитации отбора повышалась в 1,8–2 раза.
4. Далее исследования должны ответить на вопросы:
а) устойчивы ли во времени различия между семьями по химическим элементам;
б) когда хвоя приобретает эти отличия и можно ли использовать саженцы и сеянцы.
После этого опознание быстрорастущих потомств ели по химическому составу хвои (селекция по хемомаркерам хвои) вполне может оказаться прорывной технологией в селекции хвойных пород.