
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

ПАРАТГОРМОН-РОДСТВЕННЫЙ ПРОТЕИН
Курзанов А. Н., Ледванов М. Ю., Быков И. М., Медведев В. Л., Стрыгина Е. А., Бизенкова М. Н., Заболотских Н. В., Ковалев Д. В., Стукова Н. Ю.,
3.2. Существующие представления о возможных механизмах действия паратгормон-родственного белка
Поскольку большинство физиологических действий ПТГрП являются аутокринными или паракринными, а уровни его циркуляции в кровотоке очень низки, системные эффекты этого белка становится актуальным только тогда, когда происходит чрезмерное и устойчивое перепроизводство ПТГрП. Существуют только три выявленных обстоятельства, при которых ПТГрП присутствует в кровообращении в повышенных концентрациях и действует эндокринно. Это наблюдается во время беременности в период эмбрионального развития плода, когда он регулирует трансплацентарный транспорт кальция (Care A.D. et al., 1990; Grill V. et al., 1992; Wu T.L. et al., 1996; Kovacs C.S. et al., 1996; Rodda C.P. et al., 1998), в период лактации, когда этот белок, продуцируемый в молочной железе, поступает в кровоток и при различных онкологических заболеваниях сопровождающихся синдромом злокачественной гиперкальциемии при котором ПТГрП, продуцируемый опухолями, поступает в кровеносное русло (Burtis W.J. et al., 1992; Grill V. et al., 1991).
Значительно больше известно о многочисленных паракринных эффектах ПТГрП в различных тканях. Как паракринный фактор этот протеин регулирует рост и развитие многих тканей и, в первую очередь, скелета и молочных желез (McCauley L.K., Martin T.J. 2012). Паракринному действию ПТГрП отводится существенная роль в регуляции физиологического развития кератиноцитов, волосяных фолликулов, хрящей, в прорезывании зубов, регуляции функционального состояния гладких мышц, поджелудочной железы и во многих других физиологических реакциях организма. Паракринные эффекты во многом обеспечивают физиологическую регуляцию тонуса гладких мышц сосудов, кишечника, матки, мочевого пузыря, модуляцию трансэпителиального транспорта кальция в почках, плаценте, молочных железах и регулирование дифференцировки и пролиферации клеток тканей и развития органов (Martin T.J.et al., 1997; Fhilbrick W.M. et al., 1996; Naafs M., 2017). Паракринные действия ПТГрП в желудочно-кишечном тракте млекопитающих в основном связаны с релаксацией гладких мышц (Clemens T.L. et al., 2001), но сообщалось также о быстрой стимуляции кишечного транспорта Ca2+ в двенадцатиперстной кишке птиц (Zhou L.X. et al., 1992).
Внутриядерная локализация ПТГрП обеспечивает интракринные эффекты ПТГрП (Boras-Granic K. et al., 2014), которые во многом остаются неизвестными, но в целом, допускается его митогенное действие (De Miguel F. et al., 2001). Предполагается, что интракринные эффекты ПТГрП обеспечиваются фрагментом (1-87), включающим N-концевой домен и срединный фрагмент молекулы белка (midregion) (Gujral A. et al., 2001). Модуляция клеточной адгезии с участием пептида, локализованного в ядре клетки, является нормальным физиологическим действием ПТГрП, опосредованным путём увеличения транскрипции генов интегрина. Полагают, что интракринное действие ПТГрП может модулировать запрограммированную гибель клеток. Утверждается, что некоторые эффекты ПТГрП и, в частности, влияние на повышенную выживаемость хондроцитов за счет ингибирования процессов апоптоза, связаны с ядрышковой локализацией пептида (Aarts M.M. et al., 2001; Henderson J.E. et al., 1995; assfelder T. et al., 1997) (рис. 8).
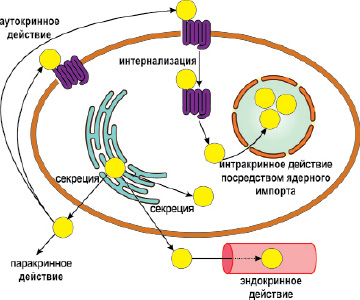
Рис. 8. Механизмы действия ПТГрП. Секретируемый ПТГрП перемещается из эндоплазматического ретикулуума сначала в цитоплазму, а затем во внеклеточное пространство, где действует паракринным или аутокринным образом посредством связывания с ПТГ/ПТГрП-рецептором. Из межклеточного пространства ПТГрП может поступть в кровоток и таким образом оказывать эндокринные влияния на клетки-мишени. ПТГрП может оставаться внутриклеточно, либо поступать в клетку путем интернализации лиганд-рецепторного комплекса и транспортируясь в ядро клетки действовать интракринно
Антиапоптотические эффекты ПТГрП выявленные в исследованиях на хондроцитах (Henderson J.E. et al., 1995) опосредованы изменением продукции антиапаптотического белка Bcl-2 (Amling M. et al., 1997). Установлено, что ядерная локализация ПТГрП играет решающую роль в регуляции экспрессии генов и, в частности, проапоптотического гена TNF, что предотвращает аноикис (специфический тип апоптоза «по умолчанию») – клеток, у которых нарушено взаимодействие с тканевым матриксом (Park S.I., McCauley L.K., 2012).