
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

ПАРАТГОРМОН-РОДСТВЕННЫЙ ПРОТЕИН
Курзанов А. Н., Ледванов М. Ю., Быков И. М., Медведев В. Л., Стрыгина Е. А., Бизенкова М. Н., Заболотских Н. В., Ковалев Д. В., Стукова Н. Ю.,
3.3.5. Роль паратгормон-родственного белка в развитии молочных желез
ПТГрП является критическим регулятором морфогенеза молочной железы у эмбрионов (Hiremath M., Wysolmerski J., 2013). Развитие эмбриональной молочной железы зависит от взаимодействий между эпителиальными и мезенхимальными клетками, которые направляют образование плакод и почек, определяют и инициируют трехмерный морфогенез, необходимый для формирования первичной протоковой системы железы (Hens J.R., Wysolmersk J.J., 2005; Veltmaat J.M. et al., 2003; Robinson G.W. et al., 1999). Молекулярные события, лежащие в основе раннего развития молочных желез, определяющие эти важнейшие эпителиально-мезенхимных взаимодействия осуществляются при участии фактора роста фибробластов, эпидермального фактора роста, сигнальных путей IHH, WNT и играют важную роль в формировании исходных плакод молочных желез (Chu E.Y. et al., 2004; Davenport T.G. et al., 2003; Hatsell S.J., Cowin P., 2006; Veltmaat J.M. et al., 2006).
Существует немало убедительных доказательств, свидетельствующих о важной роли ПТГрП в формировании молочных желез у млекопитающих (Wysolmerski J.J. et al., 1995; Boras-Granic K. et al., 2011; Gonzalez-Suarez E. et al., 2010; Wysolmerski J.J. et al., 2001; Van Houten J.N. et al., 2003). ПТГрП и PTH1R необходимы для нормального развития молочной железы. ПТГрП участвует в физиологическом развитии молочной железы (Rouffet J., Barlet J.P., 1995) являясь важным элементом организации сложного взаимодействия эпителиальных и мезенхимальных клеток (Seitz P.K. et al., 1993). Передача сигналов ПТГрП, по-видимому, имеет решающее значение для спецификации мезенхимы молочной железы, которая, в свою очередь, поддерживает эпителиальную идентичность молочной железы, направляет рост почек, нарушает рудимент мужской молочной железы и определяет образование соска (Hiremath M., Wysolmerski J., 2013). В физиологических условиях эпителиальные клетки молочных желез продуцируют много ПТГрП, секреция которого регулируется при участии рецептора кальция. Молочные железы у самок мышей с гомозиготной инактивацией гена ПТГрП не развиваются, за исключением, самых ранних этапов. Изучение плодов с мутациями в гене PTHR1, которые вызвали потерю функции рецептора подтвердило важность передачи сигналов ПТГрП для правильного развития молочной железы у людей (Wysolmerski J.J. et al., 2001). Потеря функции PTHR1 у мышей и людей приводит к полному отсутствию эпителия молочной железы (Wysolmerski J.J. et al., 1998; Dunbar M.E., Wysolmerski J.J., 1999; Foley J. et al., 2001).
Во время развития мыши ПТГрП заметно продуцируется в эпителиальных клетках молочной железы, начиная с дня E11 одновременно с образованием плакод молочной железы. PTH1R экспрессируется на незрелых мезенхимальных клетках, расположенных под всем эпидермисом. Поскольку почка молочной железы инвагинатна, ПТГрП действует на свой рецептор, чтобы вызвать дифференциацию окружающей мезенхимы в специализированную конденсированную мезенхиму молочной железы. Стимуляция ПТГрП необходима для того, чтобы эта мезенхима выполняла три жизненно важные функции:
1) поддерживать развитие эпителиальных клеток молочных желез;
2) индуцировать эпидермис для образования оболочки соска;
3) инициировать рост и морфогенез протоков.
В отсутствие сигнала ПТГрП эпителиальные клетки молочной железы дифференцируются в клетки кожи, сосок не формируется и морфогенез прерывается (Wysolmerski J.J. et al., 1998; Dunbar M.E. et al., 1999; Foley J. et al., 2001). Напротив, сверхэкспрессия ПТГрП в базальных кератиноцитах трансгенных мышей с использованием промотора кератина 14 (мыши K14-ПТГрП) приводит к превращению субэпидермальной мезенхимы из дермы в конденсированную мезенхиму молочной железы. Это, в свою очередь, подавляет развитие волосяного фолликула и заставляет эпидермис приобретать характеристики специализированной кожи сосков (Foley J. et al., 2001).
ПТГрП играет важную роль в морфогенезе молочной железы, влияя на межклеточные взаимодействия между эпителиальными клетками, формирующими протоки и альвеолы, и клетками стромы, включающими фибробласты и адипоциты. ПТГрП и его рецептор, важны для образования мезенхимы молочной железы и роста зарождающихся молочных протоков (Hens J.R., Wysolmersk J.J., 2005). Эпителиальные клетки у плода индуцируют под влиянием ПТГрП экспрессию рецепторов андрогенов в мезенхимальных клетках, которые в ответ на действие андрогенов, продуцируемых семенниками мужских эмбрионов, собираются вокруг эпителиальной почки и разрушают ее. В отсутствии ПТГрП или его рецептора в мезенхимальных клетках, в них не экспрессируются рецепторы андрогенов и из сохранной эпителиальной почки идет развитие молочной железы по женскому типу.
ПТГрП влияет на образование первичной протоковой системы железы в эмбриональном периоде (Hens J.R. et al., 2007). Удаление гена ПТГрП у мышей и людей приводит к тому, что почка молочной железы не инициирует морфогенез ветвления протоковой системы (Wysolmerski J.J. et al., 1998; Wysolmerski J.J. et al., 2001). Гиперэкспрессия ПТГрП в молочной железе трансгенных мышей сопровождается развитием дефектов формирования протоковой системы при пубертации, а также лобулоальвеолярном развитии при беременности. Мутации, вызывающие инактивацию рецептора ПТГрП, приводят к развитию летальных форм карликовости, сопровождающихся отсутствием у плодов сосков и молочных желез (Toribio R.E. et al., 2010). Продуцируемый в эпителиальных клетках эмбриональной молочной железы ПТГрП взаимодействует с мезенхимальными клетками, экспрессирующими рецептор ПТГ/ПТГрП для образования плотной мезенхимы, которая поддерживает морфогенез железы и индуцирует эпидермальные структуры к образованию соска (Wysolmerski J.J. et al., 1998).
Hens J.R., et al. (2007) изучили взаимодействие между сигналами ПТГрП и BMP во время развития ранних эмбриональных молочных желез. Авторы продемонстрировали, что сигнализация ПТГрП является разрешительной для сигнализации BMP в мезенхиме молочной железы. Показано, что ПТГрП действует, по крайней мере, частично, путем модуляции передачи сигналов BMP в клетках мезенхимы молочной железы. При образовании почек молочной железы BMP4 экспрессируется мезенхимными клетками на вентральной поверхности эмбриона. Передача сигналов ПТГрП сенсибилизирует мезенхимальные клетки к BMP in vitro и in vivo, увеличивая экспрессию рецептора BMP1A. Взаимодействие между ПТГрП и BMP4 важно для прорастания почки молочной железы. Комбинация сигналов ПТГрП и BMP увеличивает экспрессию гомеобоксного гена MSX2 в мезенхиме молочной железы, что, в свою очередь, позволяет ПТГрП подавлять образование волосяных фолликулов в зоне развивающейся почки молочной железы и соска.
Полученные данные свидетельствуют, что сигнальная передача ПТГрП активирует экспрессию BMPR1A в мезенхиме молочной железы. Это взаимодействие, в свою очередь, активирует экспрессию гена Msx2 в мезенхимальных клетках, что позволяет им подавлять образование волосяных фолликулов в коже, покрывающей соски. ПТГрП регулирует в мезенхиме молочной железы экспрессию MSX2, который является транскрипционным фактором, участвующим в эпителиально-мезенхимальных взаимодействиях во время развития. Считается, что он имеет важные функции при формировании эпидермальных придатков, поскольку нарушение гена Msx2 приводит к аномальному развитию зуба, волосяного фолликула и молочной железы (Satokata I. et al., 2000). Эти данные определяют MSX2 как важный мезенхимальный фактор, который позволяет ПТГрП и BMP4 подавлять индукцию волосяного фолликула в локусе развивающейся почки молочной железы и соска.
Сигналы ПТГрП и BMP также взаимодействуют, чтобы мезенхима могла инициировать рост эпителия почек молочной железы. Присутствие специфического BMP необходимо для полного осуществления эффектов ПТГрП. В случае нормального развития молочной железы экспрессия BMP4 в вентральной мезенхиме совпадает с образованием почек молочной железы, которые экспрессируют ПТГрП и позволяют взаимодействовать с сигнальными путями для образования мезенхимы, специфичной для молочных желез. Результаты иссдедования Hens J.R., et al. (2007) показывают, что ПТГрП-опосредованная экспрессии BMPR1A обеспечивает пространственно ограниченную, аутокринную или паракринную сигнализацию BMP внутри мезенхимы, позволяющую клеткам мезенхимы молочной железы запускать
и/или поддерживать рост эпителия зародышевой почки молочной железы.
В дополнение к стимулированию роста почек, мезенхима, специфичная для молочных желез, индуцирует формирование из эпидермиса кожи сосков, что также зависит от передачи сигналов ПТГрП. Характерной особенностью кожной оболочки сосков является отсутствие у нее волос, что, вероятно, является результатом торможения образования волосяных фолликулов ПТГрП, выделяемого эпителиальной почкой. В отсутствие ПТГрП или PTH1R волосяные фолликулы развиваются слишком близко к почке молочной железы, а в присутствии ПТГрП развитие волосяного фолликула подавляется по всему вентральному эпидермису (Foley J. et al., 2001; Wysolmerski J.J. et al., 1994). Способность ПТГрП подавлять развитие волосяного фолликула зависит от действий фактора транскрипции гомеобокс MSX2. Используя гибридизацию in situ, авторы обнаружили, что MSX2 экспрессируется в мезенхиме молочной железы (Phippard D.J. et al., 1996; Satokata I. et al., 2000). Кроме того, мезенхимальная экспрессия MSX2 требует передачи сигналов ПТГрП. Как и в молочной железе, индукция волосяного фолликула требует взаимодействий между эпителиальными и мезенхимальными клетками. Hens J.R., et al. (2007) также обнаружили, что ПТГрП и BMP4 взаимодействуют для подавления развития волос путем индуцирования Msx2 в мезенхиме молочной железы. Результаты экспериментов, представленные авторами, демонстрируют важное взаимодействие между ПТГрП и BMP4 во время развития эмбриональной почки молочной железы (рис. 10).
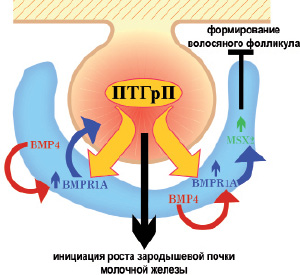
Рис. 10. Взаимодействие паратгормон-родственного протеина и костных морфогенных белков (ВМР) в инициации роста зародышевой почки молочной железы и формировании соска во время эмбрионального развития. ПТГрП секретируется из эпителиальных клеток молочных желез и повышает экспрессию рецептора ВМР (BMPR1А) в мезенхиме молочной железы. Это повышает чувствительность этих клеток к ВМР и позволяет им реагировать на BMP4 паракринно и/или аутокринно Передача сигналов ПТГрП через PTHR1 также активирует BMPR1А и сенсибилизирует мезенхимальные клетки молочной железы к BMP4, возможно, усиливая передачу сигналов BMP в мезенхиме молочной железы. Этот путь необходим для инициации роста зародышевой почки молочной железы через MMP2 и для подавления формирования волосяного фолликула в соске, индуцируя экспрессию фактора транскрипции гомеобокс Msx2, что приводит к ингибированию образования волосяных фолликулов в развивающейся оболочке соска. Передача сигналов ПТГрП также поддерживает эпителиальную идентичность молочной железы посредством передачи сигналов в мезенхиме молочной железы. В эмбриональной почке молочной железы ПТГрП оказывает влияние на морфогенез, частично, через эпителиально-стромальные перекрестные взаимодействия, опосредуемые сигналами Wnt и BMP
В лактирующей молочной железе крысы ПТГрП экспрессируется в альвеолярных эпителиальных клетках и стромальных фибробластах, а рецептор ПТГ/ПТГрП – в альвеолярных эпителиальных клетках, стромальных фибробластах и миоэпителиальных клетках (Hiremath M., Wysolmerski J., 2013). Экспрессирующиеся эпителием молочной железы RANKL и RANK-лиганд могут являться пара/аутокринными посредниками действия ПТГрП, пролактина и прогестерона. ПТГрП и пролактин повышают экспрессию RANKL. У мышей нокаунтных по RANKL или RANK имеет место недоразвитие лобулоальвеолярной системы при беременности. В современных экспериментальных исследованиях китайских ученых (Zheng H.L. et al., 2014), посвященных изучению влияния ПТГрП на транспорт кальция в молочных железах с использованием поликлональных антител против этого белка был зафиксирован уровень экспрессия ПТГрП в молочных железах значительно более высокий, чем в других органах, что свидетельствует о важной роли этого белка в регуляции функций молочных желез.