
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

ДОБЫЧА УГЛЯ В ЮЖНОЙ ЯКУТИИ И ЕЕ ВОЗДЕЙСТВИЕ НА РАСТИТЕЛЬНОСТЬ (на примере разреза «Нерюнгринский»)
Миронова С. И., Иванов В. В.,
4.1. Изменения в лишайниковом покрове
Лишайники изучены З.А. Кудиновой.
Лишайники являются неотъемлемым компонентом растительных сообществ. Изучение лишайников как индикаторов загрязнения воздуха в городах и промышленных районах имеет огромное значение в связи с проблемой охраны окружающей среды. Они считаются традиционным объектом экологического мониторинга и биоиндикации загрязнения окружающей среды.
Изучаются разные признаки лишайникового покрова, и выводы делаются на основе анализа, как всего комплекса учитываемых показателей, так и выявленных изменений только одного признака. Данные базируются на сравнительном изучении видового состава лишайников, формируемых ими группировок, распространении отдельных видов, морфологических изменений слоевищ, физиологических показателей, прироста, концентрации загрязнителей в слоевищах и другие.
Изменение видового состава лишайников. При геоботаническом исследовании территории нами выявлено 39 видов лишайников из 7 семейств, 13 родов (табл. 4.3). Ведущими семействами являются Cladoniaceae и Parmeliaceae, наибольшее количество видов представлено из рода Cladonia.
Таблица 4.3
Список лишайников окрестностей г. Нерюнгри
|
Семейство |
Род |
Вид |
|
1 |
2 |
3 |
|
Alectoriaceae (Hue) Thomas |
Alectoria Ach. |
Alectoria ochrolenuca (Hoffm.) Mass. |
|
Bryoria Brodo & D. Hawksw |
Bryoria simplicior (Vain.) Brodo & D. Hawksw. Bryoria implexa (Hoffm.) Brodo & D. Hawksw |
|
|
Cladoniaceae Zenker |
Cladonia Hill ex P. Browne |
Cladonia stellaris (Opiz.) Brodo. Cladonia amaurocraea (Florke) Schaer. Cladonia gracilis (L.) Willd. Cladonia deformis (L.) Hoffm. Cladonia uncialis (L.) Weber ex F.H. Wigg. Cladonia phyllophora Hoffm. Cladonia furcata (Huds.) Schrad. Cladonia coccifera (Florke) Spreng. Cladonia rangiferina (L.) Nyl. Cladonia arbuscula (Wallr.) Hale & W.L. Culb. Cladonia chlorophaea (Florke ex Sommerf.) Cladonia bacilliformis (Nyl.) Gluck. Cladonia mitis (Sandst.) Hustich |
|
Parmeliaceae Zenker |
Cetraria Ach. |
Cetraria cucullata (Bellardi) Ach. Cetraria laevigata Rassad. Cetraria seliaris |
|
Evernia Ach. |
Evernia esorediosa (Mull. Arg.) D.R. Evernia divaricata (L.) Ach. Evernia mesomorpha Nyl |
|
|
Hypogymnia (Nyl.) Nyl. |
Hypogymnia physodes (L.) Nyl. Hypogymnia bitteri (Lynge) Ahti |
|
|
1 |
2 |
3 |
|
Parmeliaceae Zenker |
Parmelia Ach. |
Parmelia sinuosa (Sm.) Ach. Parmelia omphalodes (L.) Ach. Parmelia centrifuga (L.) Ach. Parmelia olivaceae (L.) Ach., em Nyl. Parmelia sulcata Taylor |
|
Parmeliopsis (Nyl.) Nyl |
Parmeliopsis hyperopta (Ach.) S.L.F. Meyer. Parmeliopsis ambigua (Wulfen.) S.L.F. Meyer |
|
|
Vulpicida Mattson & M.J. Lai |
Vulpicida pinastri (Scop.) J.-E. Mattson et Lai |
|
|
Ramalinaceae C. Agardh |
Ramalina Ach. |
Ramalina sp. |
|
Stereocaulaceae Chevall. |
Stereocaulon Hoffm. |
Stereocaulon paschale Hoffm. Stereocaulon wrigthii Fuck. Stereocaulon glareosum (Savicz.) H. Magn |
|
Nephromataceae Wetm. ex J.C. David & O.E. Erikss. |
Nephroma Ach. |
Nephroma arcticum (L.) Torss. |
Распределение видов лишайников по географическим элементам и типам ареалов показало, что основу лихенофлоры окрестностей г. Нерюнгри составляет бореальный элемент, а также отмечены мультизональный, монтанный и арктоальпийский элементы. По типам ареала преобладает мультирегиональный, незначительное участие принимает голарктический тип.
При изучении изменения состава лишайников в зависимости от отдаленности от объекта загрязнения (карьера) за основу приняты результаты геохимического анализа почв (Тарабукина, 1996) и загрязнения снежного покрова (Артамонова и др., 2000; Иванов, 2005).
Нами выделены следующие зоны:
1 – зона сильного воздействия – до 5 км к юго-востоку от карьера;
2 – зона среднего воздействия – в 5–30 км от карьера;
3 – зона слабого воздействия – более 30 км от карьера.
В каждой зоне нами проводилось описание лесной растительности и сообществ эпифитных и эпигейных лишайников в 5-ти повторностях. Расстояние между площадками составляет 500 м, размер площадки 25×25 м.
Первая зона находится южнее угольного разреза «Нерюнгринский», на правом берегу р. Верхняя Нерюнгри и представляет собой пологий склон коренного берега реки.
Древостой сложен из лиственницы (Larix cajanderi) с небольшой примесью сосны и березы. Средняя высота лиственницы 20 м, средний диаметр стволов 20 см, сомкнутость крон 0,4. Подлесок слабо развит (10 %), в основном представлен березой, кедровым стлаником, душекией, в небольшом количестве встречаются можжевельник, шиповник, изредка ивы, таволга и кизильник. В травяно-кустарничковом покрове с проективным покрытием 50 %, преобладают голубика, брусника, багульник, местами произрастает шикша, из разнотравья (проективное покрытие 3–5 %) отмечены лимнас, грушанка, соссюрея, иван-чай, колокольчик, аконит, хвощ, княжек и злаки. Мохово-лишайниковый покров занимает 60 %, преобладают мхи, лишайники занимают в среднем 25 %.
Вторая зона выбрана к юго-западу от карьера на левом берегу р. Чульман, рельеф – пологий склон сопки.
Древостой состоит из лиственницы без примеси, средняя высота деревьев 20 м, средний диаметр 18 см, сомкнутость 0,3. Подлесок развит, разнопородный, проективное покрытие 30 %, преобладают береза, кедровый стланник (покрытие 20 %), также произрастают душекия, шиповник, ивы, единично отмечен курильский чай. Травяно-кустарничковый покров занимает 60–75 %, доминируют голубика, брусника, содоминируют багульник и злаки. Из трав встречаются соссюрея, грушанка, иван-чай, астрагалы, хвощ. Моховой покров почти сплошной 70 %, лишайники занимают 30–40 %.
Третья зона находится юго-западнее угольного разреза на левобережном склоне р. Чульман.
Доминирует лиственничник с примесью березы и сосны, средняя высота лиственницы составляет 20 м, диаметр – 25 см, сомкнутость крон 0,4. В составе подлеска доминирует душекия (30 %), также произрастают береза, кедровый стланик с покрытием 10–15 % и шиповник. Травяно-кустарничковый покров умеренно развит, проективное покрытие 40–50 %, произрастают голубика, брусника, багульник, из трав в небольшом количестве (5 %) лимнас, мытник, колокольчик и злаки. Мохово-лишайниковый покров разреженный, мхи занимают в среднем 30 %, лишайники – 40–50 %.
В исследованном районе выявлено 38 видов лишайников, представляющих 5 семейств, 11 родов. Наибольшее количество видов отмечено из семейств Cladoniaceae и Parmeliaceae (табл. 4.4).
Таблица 4.4
Характеристика лишайниковых сообществ на учетных площадках
|
Характеристика |
Зона 1 |
Зона 2 |
Зона 3 |
|
Общее количество лишайников |
32 |
20 |
26 |
|
Количество эпифитов |
13 |
9 |
12 |
|
Количество эпигейных видов |
17 |
10 |
14 |
|
Количество эпилитных видов |
2 |
1 |
– |
|
Среднее проективное покрытие эпигейных видов, % |
28 |
37 |
46 |
|
Среднее проективное покрытие у основания ствола, % |
0,18 |
0,37 |
0,53 |
|
Среднее проективное покрытие на высоте 1,3 м от основания ствола, % |
0,14 |
0,14 |
0,17 |
Наибольшее количество видов лишайников отмечается в сильно загрязненной зоне (зона 1), при переходе к незагрязненной (зона 3) число видов постепенно уменьшается, что можно объяснить возможным изменением рН среды от кислого до щелочной, потому что на лишайниковый покров кроме угольного разреза дополнительное отрицательное влияние оказывают сам город, автомобильный транспорт и железная дорога. Как отмечают некоторые авторы (Нильсон, Мартин, 1982), кислотность субстрата является одним из факторов, определяющих как прямое, так и косвенное влияние на жизнедеятельность лишайников. Влияние кислого загрязнения на лишайниковую флору в конечном итоге явно отрицательное, как показано большим количеством исследований. Но влияние щелочного загрязнения носит двоякий характер. С одной стороны, как при всяком загрязнении, наблюдается повреждающее действие щелочных веществ. С другой стороны, можно указать на некоторое положительное влияние щелочных веществ, особенно на фоне кислого загрязнения. Под влиянием щелочных загрязнителей наблюдается повышение рН как субстратов, так и самих лишайников. Повышение рН субстрата лишайников имеет косвенный удобрительный эффект даже в том случае, если в загрязнителе не содержится питательных веществ, необходимых для лишайников.
При переходе от сильно загрязненной зоны также увеличивается и среднее проективное покрытие эпигейных лишайников примерно в 1,5 раза. Причем у основания стволов среднее проективное покрытие увеличивается до 3-х раз, на высоте ствола 1,3 м – в 1,2 раза.
Уровни загрязнения атмосферы можно определить по индексам полеотолерантности, токсифобности, Нами по литературным источникам составлен сравнительный список видов лишайников для каждой зоны (табл. 4.5). Установлено, в районе исследований преобладают выносливые эпифитные лишайники.
Таблица 4.5
Относительная устойчивость некоторых эпифитов
к уровню загрязнения атмосферы (по литературным данным)
|
№ п/п |
Виды |
Зоны |
Субстрат |
Литературный источник (индексы полеотолерантности1, токсифобности2, уровни загрязнения атмосферы3) |
||||||||
|
1 |
2 |
3 |
4 |
С |
13 |
23 |
31 |
41 |
52 |
61 |
||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
|
1 |
Peltigera apthosa |
+ |
+ |
+ |
+ |
Г |
||||||
|
2 |
Peltigera canina |
+ |
+ |
+ |
+ |
Г |
10 |
|||||
|
3 |
Cladonia rangiferina |
+ |
+ |
+ |
+ |
Г |
||||||
|
4 |
Cetraria cucullata |
+ |
+ |
+ |
+ |
Г |
||||||
|
5 |
Cetraria laevigata |
+ |
+ |
+ |
+ |
Г |
||||||
|
6 |
Parmeliopsis hyperopta |
+ |
+ |
+ |
+ |
Ф |
3 |
3 |
4, 8, 9 |
1,2 |
||
|
7 |
Evernia mesomorpha |
+ |
+ |
+ |
+ |
Ф |
1,2 |
1–3 |
5 |
10 |
1–6 |
|
|
8 |
Parmelia sinuosa |
+ |
+ |
+ |
+ |
Ф |
1 |
10 |
||||
|
9 |
Hypogymnia physodes |
+ |
+ |
+ |
+ |
Ф |
2,4 |
1,2 |
1–3 |
6 |
8 |
4 |
|
10 |
Bryoria simplicolor |
+ |
+ |
+ |
+ |
Ф |
2,3 |
|||||
|
11 |
Bryoria implexa |
+ |
+ |
+ |
+ |
Ф |
2,3 |
4 |
7 |
|||
|
12 |
Vulpicida pinastri |
+ |
+ |
+ |
+ |
Ф |
3 |
3 |
3 |
|||
|
13 |
Cladonia stellaris |
+ |
+ |
+ |
+ |
Г |
||||||
|
14 |
Cladonia amaurocraea |
+ |
+ |
+ |
+ |
Г |
||||||
|
15 |
Cladonia gracilis |
+ |
+ |
+ |
+ |
Г |
||||||
|
16 |
Stereocaulon paschale |
+ |
+ |
+ |
Г |
|||||||
|
17 |
Hypogymnia bitteri |
+ |
+ |
+ |
Ф |
7 |
1 |
|||||
|
18 |
Cladonia deformis |
+ |
+ |
+ |
Г |
|||||||
|
19 |
Cladonia uncialis |
+ |
+ |
+ |
Г |
|||||||
|
20 |
Stereocaulon glareosum |
+ |
+ |
Г |
||||||||
|
21 |
Parmelia sulcata |
+ |
+ |
Ф |
2–4 |
1–5 |
1–3 |
5, 7 |
1–9 |
5,6 |
||
|
22 |
Cladonia phyllophora |
+ |
+ |
Г |
||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
|
23 |
Parmelia olivaceae |
+ |
+ |
Ф |
5 |
2,6 |
1–5 |
|||||
|
24 |
Stereocaulon wrigthii |
+ |
+ |
Л |
||||||||
|
25 |
Parmelia omphalodes |
+ |
+ |
Ф |
8,5 |
5 |
||||||
|
26 |
Parmelia centrifuga |
+ |
Ф |
|||||||||
|
27 |
Cladonia furcata |
+ |
Г |
|||||||||
|
28 |
Cladonia coccifera |
+ |
Г |
|||||||||
|
29 |
Nephroma arcticum |
+ |
Л |
|||||||||
|
30 |
Cladonia arbuscula |
+ |
Г |
|||||||||
|
31 |
Cetraria seliaris |
+ |
Г |
|||||||||
|
32 |
Evernia esorediosa |
+ |
Ф |
1–3 |
1–6 |
|||||||
|
33 |
Parmeliopsis ambigua |
+ |
+ |
+ |
Ф |
3 |
1,2 |
4, 5 |
1–9 |
2,3 |
||
|
34 |
Cladonia chlorophaea |
+ |
+ |
Г |
4 |
1–3 |
||||||
|
35 |
Cladonia bacillaris |
+ |
Г |
|||||||||
|
36 |
Cladonia mitis |
+ |
Г |
|||||||||
|
37 |
Evernia divaricata |
+ |
Ф |
1–3 |
2 |
9 |
1,2 |
|||||
|
38 |
Alectoria ochrolenuca |
+ |
Ф |
|||||||||
Примечания: Субстрат произрастания, обозначения: Г – эпигейный вид, Ф – эпифит, Л – эпилит. Литературный источник: 1 – Журавлева, 2005, 2 – Меркулова, 2006, 3 – Баумгертнер, 1999, 4 – Трасс, 1985, 5 – Инсарова, Инсаров, 1989, 6 – Рыкова, 2004.
Изменение биохимического состава лишайников
Воздействие открытой добычи угля на растительность можно дополнить и изменением биохимического состава лишайников.
В любом живом организме постоянно протекают реакции окисления субстрата, являясь факторами регуляции важнейших метаболических процессов. Постоянство уровня суммарной антиокислительной активности тканей и индивидуальность этого уровня для каждого организма служат одним из основных показателей гомеостаза. При негативном для организма влиянии окружающей среды, изменении определенных факторов происходит ответная защитная реакция на раздражитель. В первую очередь резко активизируются процессы антиоксидантной защиты от свободнорадикального окисления субстратов (Журавлева, 2005).
Биологический механизм биоантиокислителей сводится к смещению конкурентного отношения свободнорадикального и ферментативного окисления в пользу ферментативного, тем самым биоантиокислители регулируют степень подавляющего влияния свободнорадикального окисления на большинство метаболических процессов. Конечным итогом действия биоантиокислителей является создание оптимальных условий для метаболизма и обеспечение нормального роста клеток и тканей (Баумгертнер, 1999). В любом живом организме постоянно протекают реакции окисления субстрата, являясь факторами регуляции важнейших метаболических процессов. Постоянство уровня суммарной антиокислительной активности тканей и индивидуальность этого уровня для каждого организма служат одним из основных показателей гомеостаза.
В экстремальных неблагоприятных условиях окислительные процессы в клетке активируются, расход антиоксидантов увеличивается и, когда концентрация последнего падает ниже критической, окисление липидов резко возрастает. Окисление липидов приводит к разрушению комплекса белок-липид, патологии мембран и гибели клетки (Инсарова, Инсаров, 1989). Для преодоления этого воздействия организмы выработали комплексную, многоступенчатую превентивную систему, состоящую из многочисленных ферментов, таких как супероксиддисмутаза (СОД), каталаза, глутатион пероксидаза и др., а также из низкомолекулярных антиоксидантов (НМАО).
В качестве объекта исследования нами выбраны широко распространенные на территории Якутии лишайники Cladina stellaris и Cetraria laevigata (Соловьева, Кудинова, 2007). Отбор проб проводился в конце июля и в начале августа. В качестве контроля выбрана зона 3.
Активность антиоксидантных систем (kаоз) лишайников и коэффициент антиоксидантно-прооксидантного равновесия (kаоз / kПОЛ) рассчитывали по формулам:
(kаоз)N = S(параметры АО-систем)N / кол-во изученных АО-систем,
где (параметры АО-систем)N – нормированные к контролю значения НМАО, СОД;
(kаоз / kПОЛ)N = (kаоз)N / [активность ПОЛ]N,
где активность ПОЛ N – нормированное к контролю содержание МДА.
Суммарное содержание НМАО и фермента СОД, у обеих видов лишайников увеличивается с переходом от более загрязненной зоны (зона 1) к менее загрязненной зоне (зона 3), при этом по отношению к контрольной зоне содержание НМАО повышено, а активность СОД понижена.
Процессы перекисного окисления липидов у двух видов лишайников почти в 3 раза интенсивнее протекают в зоне 1, по сравнению с фоновой зоной (рис. 4.1, а и б). При этом у вида Cetraria laevigata при переходе в зоны 2 и 3 процессы ПОЛ менее выражены, что объясняется высокой активностью защитных систем в слоевище лишайника.
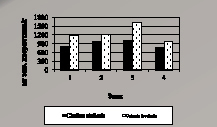
Рис. 4.1, а. Суммарное содержание НМАО
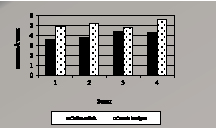
Рис. 4.1, б. Активность фермента
в лишайниках мг экв. кверцетина/г СОД мкмоль/г мин
В загрязненной зоне 1, при наибольшей пылевой нагрузке, происходит истощение антиоксидантной системы лишайников, вследствие этого резко возросло окисление липидов, которое со временем приводит к разрушению комплекса белок-липид, патологии мембран и гибели клетки. Наиболее резко защитные механизмы активизированы в зоне 3, где отмечается сравнительно наибольшее содержание антиоксидантов в лишайниках (табл. 4.6).
Таблица 4.6
Показатели антиоксидантной защиты лишайников
по отношению к контролю
|
Вид лишайника |
Зоны техногенной нагрузки |
Σ НМАО |
Активность СОД |
ПОЛ |
kаоз |
k аоз/пол |
|
Cladonia stellaris |
1 |
1,07 |
0,84 |
2,96 |
0,96 |
0,32 |
|
2 |
1,29 |
0,89 |
2,18 |
1,09 |
0,5 |
|
|
3 |
1,35 |
1,03 |
1,62 |
1,19 |
0,73 |
|
|
4 контроль |
1 |
1 |
1 |
1 |
1 |
|
|
Сetraria laevigata |
1 |
1,19 |
0,89 |
2,73 |
1,04 |
0,38 |
|
2 |
1,24 |
0,92 |
1,06 |
1,08 |
1,02 |
|
|
3 |
1,66 |
0,87 |
1,08 |
1,27 |
1,18 |
|
|
4 контроль |
1 |
1 |
1 |
1 |
1 |
В лишайниках Cladina stellaris и Cetraria laevigata коэффициент антиоксидантной защиты и коэффициент антиоксидантно-прооксидантного равновесия смещен в сторону усиления защитных реакций при переходе от зоны 1 к зоне 3. Как видно из показателей kаоз / kПОЛ равновесие систем сильно нарушено и смещено в сторону ПОЛ у вида Cladina stellaris. Вид Cetraria laevigata оказался наиболее устойчивым к неблагоприятным факторам окружающей среды, коэффициент равновесия смещен в сторону усиления антиоксидантной защиты.
Таким образом выявлено, что лишайники вида Cladina stellaris, более чувствительны к изменениям в окружающей среде, чем лишайники вида Cetraria laevigata. Интенсивность процессов перекисного окисления липидов и уровни содержания антиоксидантов показывают степень угнетения, ответ организма на неблагоприятные воздействия окружающей среды. Конечным результатом описанных выше процессов является угнетение таких важных процессов жизнедеятельности как рост и развитие организма талломов лишайников в результате патологии и гибели клеток.