
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

3.3. Реакции живого организмов на действие холода
В настоящее время наукой не определено существование границы между теплом и холодом, что предполагает отсутствие ответной реакцииживого организма на действие холода. В этом случае, кривая кинетики затухания процессов жизнедеятельности по мере падения температуры должна подчиняться известному правилу Вант-Гоффа и иметь линейный характер. Предполагается, что физиолого-биохимические процессы останавливаются в результате замерзания воды в клетках и тканях живого организма.
Однако, имеющиеся некоторые эмпирические и экспериментальные материалы, дают возможность предполагать наличие более или менее четкой границы между теплом и холодом, которая расположена несколько выше 0 °С Цельсия. Как показывают наблюдения на природе и результаты опытов, жизнедеятельность живых организмов довольно резко подавляется при более высокой температуре, чем 0 °С, когда вода еще находится в жидком состоянии. Это может быть обусловлено существованием фактора, отличного от льда, ограничивающего обмен веществ у живых организмов в условиях пониженных температур.
Г. Селье [253], автор открытия явления стресса у живых организмов, назвал стресс неспецифической реакцией организма на любые раздражители. Стресс (от англ. stress – давление, нажим, напор; гнет; нагрузка; напряжение) – это интегральный неспецифический ответ организма на повреждающее действие, направленный на выживание организма за счет мобилизации и формирования защитных систем. Такой сильнодействующий фактор внешней среды, способный вызвать в организме повреждение или даже привести к смерти, называют стрессорным фактором, или стрессором. Стрессоры подразделяются на три основные группы: физические, химическеие и биологические. К физическим стрессорам относятся: недостаточная или избыточная влажность, освещенность, температура, радиоактивное излучение, механические воздействия, аноксия и т. д. Несмотря на то, что холод сам по себе и механизм его действия на физиолого-биохимические процессы, по существу, неизвестны, но его отрицательное действие на живые системы было доказано практикой и экспериментальными исследованиями. Таким образом, холод, как неблагоприятный фактор, должен выступать как стрессор. Стресс, как известно, вызывает ответную реакцию [215].
Для определения наличия реальной границы между теплом и холодом нужны были методы, позволяющие зарегистрировать ответную реакцию организма на холод. В этом плане, самым подходящим был метод непрерывного слежения за кинетикой активности физиологических процессов у живых организмов. Стрессовую реакцию организма можно уловить при помощи приборов, регистрирующих кинетику основных физиологических процессов, таких как дыхание, фотосинтез, биоэлектрическая активность и др.
Как показали результаты наших исследований и материалы из литературных источников, в кинетике интенсивности физиологических процессов на органоидном, клеточном, тканевом и организменном уровнях, наблюдается аномальные изменения в области +4 °С. Приведем из них несколько характерных примеров.
Очень удобным тест-методом для массового определения реакции растений на действие холода оказался метод непрерывной регистрации биоэлектрических потенциалов. Ответную биоэлектрическую реакцию на раздражитель-стрессор в своих опытах с лягушками впервые наблюдал основоположник электрофизиологии итальянский ученый Л. Гальвани. Метод непрерывной регистрации ответной реакции тканей животных и растений успешно применял в своих опытах Ч. Бос [41].
Нами в опытах был применен известный метод И.И. Гунара [95, 96], с незначительной модификацией. Нам важно было регистрировать характерный излом в кривой кинетики величины биопотенциалов по мере охлаждения органов или тканей растений, который обычно наблюдается при действии какого-либо раздражителя-стрессора [223].
Изучалось большое количество дикорастущих и культурных растений, относящихся к различным семействам в том числе из семейства Бобовых (Leguminosae Juss) – клевер лупиновидный (Trifolium lupinaster L.), вика мышиная (Vicia cracca L.), из; Астроцветных (Asteraceae) – одуванчик рогоносный (Taraxacum ceratoforum (Ledeb.) DC), из Лютиковых (Ranunculaceae Juss.) – прострел желтеющий (Pulsatilla flavescens Juss.), из Подорожниковых (Plantaginaceae Juss.) – подорожник большой (Plantago major L.), из Мятликовых (Poaceae) – пырей ползучий (Elytrigia repens L.), из Розоцветных (Rosacea Juss.) – кровохлебка аптечная (Sanguisorba officinalis L.), а также древесно-кустарниковые – лиственница Каяндера (Larix cajanderi Mayr), береза плосколистная (Betula platyphylla Sukacz.), ива сухолюбивая (Salix bebbiana Sarg.), культурные растения – пшеница сорта «Якутянка», огурцы сорта «Парниковые» и др. [303].
Оказалось, что растения, растущие в нормальных условиях среды, на действие холода активно не реагируют, то есть кривая кинетики биопотенциалов имеет спокойный характер. Совсем другая картина наблюдается после холодового закаливания растений. Так, закаленные холодом холодостойкие растения на действие холода реагируют резким скачком величины биопотенциалов в области +4 °С.
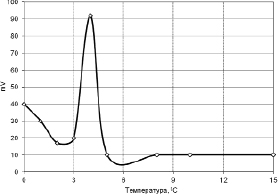
Рис. 3.3.1. Кинетика БЭП листьев пшеицы при охлаждении [300]
На рис. 3.3.1 приведена кривая кинетики биопотенциалов пшеницы сорта «Якутянка» по мере ее охлаждения, где виден резкий скачок величины биопотенциалов в области +4 °С. Такой скачок величины биопотенциалов характерен и у других исследованных растений после их холодового закаливания [300]. Как известно, биоэлектрические потенциалы непосредственно связаны с процессами жизнедеятельности клеток и отражают интенсивность протекающих в них процессов обмена веществ.
Фотохимическую активность (ФХА) по методу восстановлению феррицианида калия [68, 69] изучали у хлоропластов, выделенных из листьев пшеницы сорта «Якутянка» и огурца сорта «Парниковые».
Как видно на рис. 3.3.2 по мере падения температуры фотохимическая активность хлоропластов огурца и пшеницы до 6 °С снижается более или менее равномерно, начиная с 5 °С, резко падает и при температуре +4 °С и ниже при 5-минутной экспозиции восстановление феррицианида калия не происходит [305]. Как известно, пшеница является холодостойким растением, а огурцы относсится к теплолюбивым. Интересно, что несмотря на такое существенное различие по отношению к теплу, реакция у этих двух растений на действие температуры ниже +4 °С имеет одинаковый характер.
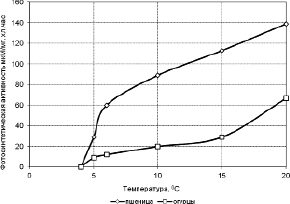
Рис. 3.3.2. Кинетика фотохимической активности (ФХА) изолированных хлоропластов огурцов и пшеницы при охлаждении [305]
Такая же картина наблюдается в кинетике интенсивности дыхательной активности (ДХА) изолированных митохондрий у указанных растений. Дыхательная активность митохондрий в процессе их охлаждения постепенно подавляется при снижении температуры с 20 °С до 5 °С и кривая падения ДХА имеет почти линейный характер.
Интересно отметить, что митохондрии все еще довольно активно функционирующие при температуре +5 °С при снижении температуры всего на 1 °С, т. е. при температуре +4 °С, практически инактивируются – во всяком случае, в течение 5 минут, за которые инкубировались митохондрии в дыхательном субстрате при изучаемой температуре, поглощение кислорода митохондриями обнаружено не было (рис. 3.3).
Таким образом, нижней точкой или границей фотохимической активности изолированных хлоропластов и активной дыхательной деятельности митохондрий теплолюбивых и холодостойких растений является одна и та же температура – +4 °С.
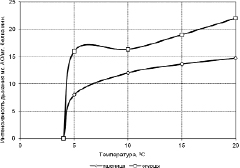
Рис. 3.3.3. Кинетика дыхательной активности (ДХА) изолированных митохондрий огурцов и пшеницы при охлаждении [305]
Резкое падение интенсивности энергетического обмена у органоидов растений при температурах ниже +4 °С можно интерпретировать как криошок, вызываемый действием холода, что предполагает наличие такой же вспышки физиологических процессов у закаленных в холоде растений.
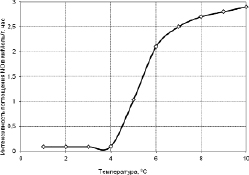
Рис. 3.3.4. При температурах ниже +4 °С незакаленными активновегетирующими растениями пшеницы поглощение ионов NO3 не происходит (криошок) [314]
Дальнейшие наши исследования показали, что действительно это так. На рис. 3.3.4 мы видим, что при температурах ниже +4 °С незакаленными активновегетирующими растениями пшеницы поглощение ионов NO3 не происходит (криошок). А у прошедших холодовое закаливание растений пшеницы, наоборот, наблюдали значительное усиление поглощения этого иона при пониженных температурах, начиная с +4 °С (рис. 3.3.5).
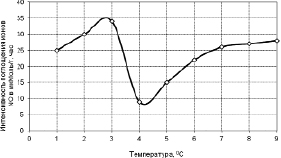
Рис. 3.3.5. Кинетика поглощения ионов, закаленных холодом растений. Наблюдается усиление поглощения NO3, начиная с +4 °С [314]
Среди физиолого-биохимических факторов защиты растений от стрессовых воздействий пероксидазу рассматривают как одну из важнейших каталитических систем, активно участвующих в авторегуляции метаболизма при стрессе. Влияние экстремальных факторов среды, в том числе низких температур, приводит к активации пероксидазы в различных тканях и органах живых систем. Установлено, что при температурах, не убивающих клетку, возможны процессы, способствующие восстановлению нарушенных структур и функций.
В связи этим нами также изучена активность пероксидазы в проростках гороха сорта «Капитал». В опытах были получены аналогичные предыдущим данным результаты. Так в тканях незакаленных растений гороха содержание пероксидазы с понижением температуры постепенно падает до температуры около +4 °С и при дальнейшем снижении температуры количество ее практически не меняется (рис. 3.3.6). В тканях, закаленных холодом растений, наоборот, наблюдается скачкообразный пик содержания пероксидазы в области +4 °С (рис. 3.3.7).
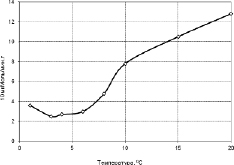
Рис. 3.3.6. Активность перооксидазы в тканях проростков гороха до закаливания [314]
По литературным данным при холодовом закаливании состав пероксидазы существенно меняется – появляются новые структуры, представляющие собой изомерами пероксидазы, устойчивые и активные в условиях холода. Как известно, данный фермент входит в состав антиоксидантной системы растений, способен катализировать реакции оксидазного, пероксидазного и оксигеназного окисления.
Субстратами пероксидазы могут быть неорганические и органические соединения. Было показано, что в реакциях совместного окисления, в присутствии двух и более субстратов, наблюдаются эффекты их взаимного влияния на каталитический процесс.
Негативное влияние холода зависит от диапазона понижения температур и продолжительности их воздействия. Уже неэкстремальные низкие температуры неблагоприятно сказываются на растениях, поскольку тормозят основные физиологические процессы (фотосинтез, транспирацию, водообмен и т. д.), снижают энергетическую эффективность дыхания, изменяют функциональную активность мембран, приводят к преобладанию в обмене веществ гидролитических реакций.
Как показали иследования интенсивнолсти дыхания клубней картофеля, подавление этого процесса холодом идет только до +4 °С, а при дальнейшем снижении температуры интенсивность дыхания усиливается. При этом энергетическая эффективность дыхания снижается, только незначительная часть энергии дыхания фиксируется в АТФ, а ее львиная доля выделяется в виде тепла. Поэтому наиболее оптимальной температурой хранения клубней картофеля является температура +4 °С. По данным Дж. Нэш [202] минимальный уровень интенсивности дыхания у клубней наблюдается в области +4 °С (рис. 3.3.8).
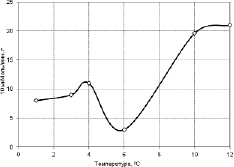
Рис. 3.3.7. Активность пероксидаз в тканях из проростков гороха после закаливания [314]
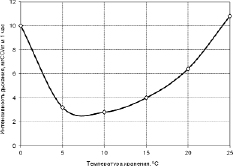
Рис. 3.3.8. Кинетика инстенсивности дыхания клубней картофеля при охлаждении [202]
В опытах Wassink [401] в листьях подсолнечника при пониженных температурах гидролиз крахмала наиболее интенсивно идет в области +4 °С.
Н.И. Николаева и др. [199] исследовали яровизацию семян различных культур растений при разных температурах и установили, что наиболее благоприятной температурой для яровизации растений является температура, близкая к +4 °С. На рис. 3.3.9 приводятся данные об яровизации у японской редьки, которая показывает, что она идет с максимальной интенсивностью при температуре близкой к +4 °С, а при более низких температурах этот процесс практически приостанавливается.
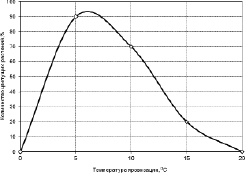
Рис. 3.3.9. Яровизация японской редьки при различных температурах (по Kamura, 1961 цит. по [199])
Как пишет Н.И. Калабухов [135], температура тела у зимующего суслика держится на уровне близкой к +4 °С.
Очень интересные данные по выходу гемоглобина из эритроцитов и об устойчивости мерцательного эпителия и диафрагмальной мышцы крысы, хомяка и суслика (в активном состоянии и в период зимовки) к охлаждению мы находим в работе Л.Г. Вольфензон и др. [66].
Как видно на рис. 3.3.10 кинетика выхода гемоглобина у гетеротермного суслика и гомойотермной крысы при различных температурах имеет одинаковый характер. Так, как у суслика, так и у крысы выход гемоглобина из эритроцитов усиливается при температурах ниже +4 °С, которая является переломной.
Аномалии в области температуры +4 °С встречаются также в кривой кинетики пики устойчивости мерцательного эпители и диафрагмальной мышцы крысы, хомяка и суслика к охлаждению (рис. 3.3.11).
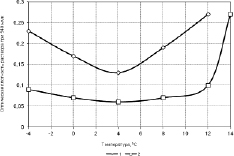
Рис. 3.3.10. Выход гемолглобина из эритроцитов активного суслика (2) и крысы (1) после 25 часов инкубаци при различных температурах [66]
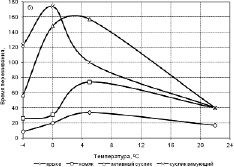
Рис. 3.3.11. Устойчивость мерцательного эпителия (а) и диафрагмальной мышцы (б) крысы, хомяка и суслика (активного и зимующего) к охлаждению [66]
А.И. Дащковским [102] определялось содержание кислорода в ткани мозга лягушки при охлаждении и показано, что концентрация кислорода в тканях в основном зависит от следующих факторов:
1) скорости мозгового кровотока и транспортных свойств крови;
2) интенсивности утилизации кислорода тканью мозга;
3) растворимости кислорода в плазме крови и тканевой жидкости.
При понижении температуры первый фактор может обусловливать снижение кислорода в ткани, а второй и третий – повышение. В опытах в качестве исходного фона регистрировалась содержание кислорода в мозгу лягушки при вдыхании воздуха при комнатной температуре (20–23 °С).
В опытах лягушки переводились на дыхание чистым кислородом. Для оценки изменения содержания кислорода в тканях мозга лягушек в процессе охлаждения за 100 % принимались значения показателя диффузионного тока при температуре 2 °С. Относительные показатели кислорода в мозгу при различных температурных условиях показаны на рис. 3.3.12, где для сравнения характеристики изобарных концентраций кислорода для воды в зависимости от растворимости при разных температурах.
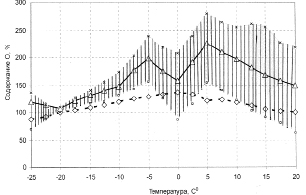
Рис. 3.3.12. Кривая содержания кислорода в тканях мозга лягушек при охлаждении и последующем согревании [102]
Из приведенного материала следует, что концентрация кислорода в мозгу лягушек по мере его охлаждения неизменно растет вплоть до 5 °С. Это объясняется преоблаждающим снижения интенсивности потребления кислорода тканью по сравнению со снижением скорости кровотока.
При согревании лягушек кривая приобретает противоположное направление и в пределах вариабельности приближается к исходному уровню, взятому при 2 °С. Концентрация кислорода при –0 °С достоверно ниже уровня, полученного при 5 °С и в период охлаждения и согревания.
Таким образом интенсивность утилизации кислорода мозгом лягушки при температурах ниже 5 °С существенно ускоряется.
Усиление интенсивности дыхания при околонулевых температурах наблюдается и у зимоспящих теплокровных животных. Оно имеет огромное значение для животных для избежания явной смерти от замерзания.
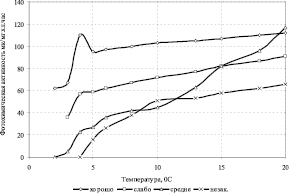
Рис. 3.3.13. Влияние условий содержания растений на кинетику ФХА хлоропластов при охлаждении на примере пшеницы сорта «Якутянка-224» [308]
Н.И. Калабухов [134] приводит данные опытов Кайзера, которым было определено дыхание сусликов при различных температурах. Им установлено, что суслики в спячке потребляли при 5 °С в течние часа на 1 кг веса 19,7 мл кислорода. При понижении температуры до 2 °С у них наблюдался рост этого показателя почти в 2 раза – 38,9 мл. В результате такого усиления дыхания животные пробуждались.
Таким образом, явление стимуляции дыхательного процесса при низких околонулевых положительных температурах наблюдается и у растительных и животных организмов. Это закономерно, так как на однотипность реакций растительной и животной клетки на действие внешних стрессорных факторов указал еще Н.Е. Введениский [54]. Интерес но здесь то, что перелом в кинетике дыхания происходит как и в динамике других физиологических процессов в подобных условиях, в области +4 °С, на что раньше авторы внимания не уделяли.
Реакция живых организмов на действие пониженной температуры зависит от предшествующих условий содержания и от отношения организма к температурному фактору (рис. 3.3.13). Применительно к растениям, как считает В.В. Полевой [228] можно говорить о следующих трех фазах стрессовой реакции:
1) первичной стрессовой реакции;
2) адаптации;
3) истощения ресурсов надежности.
Первичная стрессовая реакция в наших опытах наблюдается у активно растущих растений в виде криошока – все физиологические процессы как бы замирают. Эта реакция вызывается холодом, начиная с +4 °С. Вторая фаза, как мы считаем, состоит из нескольких подфаз или стадий непрерывно и последовательно протекающего адаптационного процесса. В подфазе репарации растение оправляется от криошока и начинается возобновление физиологических функций. Во второй стресс-синтетической подфазе, возобновившиеся после криошока, физиологические процессы существенно усиливаются, достигая иногда величин интенсивности этих процессов при температурах 15 и 20 °С. Эти процессы идут ниже 5 °С, начиная с 4 °С. Об этом свидетельствуют установленные нами в области +4 °С резкие переломы хода физиолого-биохимических процессов в сторону их усиления. Это указывает на то, что они качественно отличаются от подобных же процессов, протекающих при нормальных температурных условиях. Действительно, в этих условиях идет синтез стресс-белков, гидролиз сложных макромолекул и накопление веществ, выполняющих роль криопротекторов и т. д. [64] и др.
Вторая фаза длится 20–30 дней и в результате формируется стресс-структуры в мембранах, цитоплазме, митохондриях и ядре и клетка приобретает новую молекулярно-биохимическую стресс-архитектонику, устойчивую к холоду.
Третья подфаза – это собственно состояние приспособления, или адаптированное состояние, когда достигнута устойчивость, адекватная силе воздействующегося холода. При этом надежность системы функционирования растительного организма существенно повышается [91]. В это время физиологические процессы сильно замедляются и растение постепенно переходит в состояние вынужденного покоя.
Наличие фазы истощения для холодоустойчивых растений может быть дискуссионным и представляет теоретический интерес, поскольку переход в состояние покоя предотвращает истощение растительного организма, тем самым выжить в неблагоприятных условиях существования. У теплолюбивых растений при действии холода наоборот ярко выражены первая и четвертая фазы, а вторая фаза и фаза адаптации, протекает очень слабо или отсутствует.
Очевидно, температуры ниже 5 °С, то есть, начиная с 4 °С, являются стрессорными, действие которых вызывает стрессорную реакцию, соответствующую первой фазе холодового закаливания. Другими словами, холод, как неблагоприятный экологический фактор на уровне физиологических процессов начинается с температуры +4 °С.
Таким образом, растения в активно растущем состоянии дают четкую стресс-реакцию в виде криошока, а во второй стресс-синтетической подфазе – усиление физиологических процессов, во время которого идет усиленный стресс-синтез биологических соединений, устойчивых к холоду.
В большинстве случаев исследователи в своих экспериментах имеют дело с растениями, находящимися в репарационной подфазе адаптации к холоду, и такое слабое закаливание растений происходит во время подготовки к экспериментам и в ходе самих экспериментов, которые проводятся в холодных условиях. Следует иметь в виду также, что растения в силу ряда причин могут находиться в состоянии неспецифической устойчивости. Такие растения становятся толерантными к действию холода и не дают четкую стресс-реакцию на холод. Это наблюдение может иметь методическое значение и учтено при проведении физиологических исследований.
Практически для того, чтобы наблюдать хорошо выраженную реакцию организма, в частности, растительного на действие температур, лежащих в области +4 °С и ниже, необходимо брать активновегетирующие экземпляры, растущие в самых благоприятных условиях или растения, прошедшие 1–2 фазу холодового закаливания.
Как известно, скорость биохимических реакций у пойкилотермных организмов зависит от температуры среды. Как видно из приведенных выше материалов, правило Вант-Гоффа более или менее приемлемо при понижеии температуры только до +4 °С, а ниже этой температуры ход физиологических процессов и у растений и у животных организмов аномально меняется. Это можно расценивать, как ответную стресс-реакцию живых организмов на неблагоприятное действие пониженной температуры, то есть на действие холода [302, 308].
Как было указано выше, холод является стрессором или неблагоприятным фактором для протекания метаболических процессов, а тепло, наоборот, способствует нормальной жизнедеятельности организмов, то есть теплые температуры можно рассматривать как положительные, а холодные – как отрицательные.
Таким образом, можно утверждать, что температурная граница между теплом и холодом объективно находится при температуре +4 °С.
После установления границы между холодом и теплом, нам представляется весьма интересным вопрос выяснения причины аномального поведения физиологических процессов, начинающихся с +4 °С или причины неблагоприятного действия холода на организм. К выяснению этого вопроса посвящена следующая глава.