
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

8.2. Биологический нуль как общебиологическое понятие
В живом организме одновременно идут процессы ассимиляции и диссимиляции, преобладание одного из которых определяет в данный момент состояние организма в целом. Если более интенсивно идет ассимиляция – организм будет в активном состоянии, будет расти и развиваться. В случае преобладания диссимиляции над ассимиляцией организм переходит в другое физиологическое состояние, характеризующееся пассивностью. Указанные процессы зависят от температуры, причем с понижением температуры гидролитические процессы активизируются. Это обусловлено, как мы уже знаем, преобладанием в клетке льдоподобной формы воды. При температурах, начиная с 4 °С и ниже, в воде теоретически исчезает плотноупакованная вода, ответственная за синтетические процессы. Однако было бы рискованным утверждать, что при этих температурах, например, у всех растений, полностью прекращаются синтетические процессы, поскольку реакция растений на действие холода различна, и зависит от терморезистентности растений. В связи с этим мы рассмотрим влияние температур, лежащих ниже 4 °С для каждой группы растений в отдельности.
В настоящее время растения делят на 3 большие группы:
1. Теплолюбивые – растения тропиков или сельскохозяйственные культуры тропического происхождения, погибающие в условиях холода.
2. Холодоустойчивые – растения южного происхождения, выдерживают холод, но погибают при небольшом морозе.
3. Морозоустойчивые – типичные растения средних и северных широт, выдерживающие замораживание тканей.
Как мы знаем, теплолюбы имеют ликвофильный тип водообмена и являются типичными гласифобами. В связи с этим, нижней границей жизни у истинных теплолюбов является температура 4 °С, когда вода приобретает, в основном, льдоподобную структуру, и при этой температуре полностью инактивируется теплолюбивый архекомплекс систем жизнедеятельности (АСЖД). Ассимиляция практически равна нулю.
Холодоустойчивые растения имеют ликвогласифильный тип водообмена и при температуре 4 °С практически прекращают свою синтетическую деятельность в связи с потерей функции архекомплекса систем жизнедеятельности, однако они не погибают, благодаря функционированию системы гипобиометаболизма (СГБМ), работоспособной в среде льдоподобной воды. В этих условиях процессы ассимиляции практически прекращаются. Типичным примером является клубень картофеля. Клубни картофеля не прорастают при температурах, лежащих ниже 4 °С. Они также после закаливания не становятся невымерзающими [120, 308].
Температура ниже 4 °С резко подавляет синтетические процессы и у морозоустойчивых растений. Это также связано с потерей активности архекомплекса систем жизнедеятельности. Второй же комплекс систем жизнеобеспечения и синтеза у этих растений, связанный в своей деятельности с льдоподобной водой, активно включается также при 4 °С и более совершенен, чем у холодоустойчивых растений. В результате в этих условиях синтезируются криобелки, растворимые углеводы, непредельные жиры и другие криорезистентные структуры клетки, способствующие морозостойкость растения. В начальных этапах действия холода ассимиляция может незначительно преобладать над диссимиляцией, однако со временем она постепенно затухает.
Таким образом, у различных групп растений, в зависимости от их холодоустойчивости, при температурах ниже 4 °С синтетические процессы прекращаются вообще или резко подавляются, или начинают кратковременно активизироваться в начальный период адаптации.
Другими словами, температура 4 °С является той важной и единственной температурой, при которой происходят качественные изменения обмена веществ и +4 °С вполне справедливо может называться биологическим нулем у растений.
А как обстоят дела у других организмов? Как было показано в предыдущих главах, аномалия физиологических процессов в области 4 °С встречается у самых разных представителей растительного и животного царств, царства грибов – у низших, простейших и высокоорганизованных, стоящих на высшей ступени эволюционного развития, что в общем виде показано в табл. 8.2.1.
Таблица 8.2.1
Аномальная реакция разных представителей живого мира на действие температуры +4 °С
|
Систематическое положение |
Некоторые представители |
Источники |
|
Царство животных Беспозвоночные Одноклеточные животные |
Тип простейшие Инфузории, амебы |
Полянский, Суханова [229]; Суханова [280] |
|
Тип мягкотелые (моллюски) |
Littorina littorea |
Newel [379] |
|
Класс насекомые |
Шелкопряд, слепни, ручейники |
Дре [108]; Васюкова [52]; Каширская [139] |
|
Позвоночные Тип хордовые Класс рыбы |
Хамса, сельд, караси и др. |
Шмидт [335]; Моисеев и [188]; Кириллов [141] |
|
Класс земневождные |
Лягушки |
Дашковский [102]; Наумов [194]; Пегель [217] |
|
Класс пресмыкающиеся |
Живородящие |
Седалищев, Белимов [251] |
|
Класс млекопитающие а) Зимоспящие |
Сурки, суслики, полчки и др. |
Бибиков [30]; Калабухов [135]; Соломонов и др. [267]; Ануфриев [18] |
|
б) Незимеспящие |
Крысы и др. |
Вольфензон и др. [66]; Майстрах, [168]; Штарк [337] |
|
Царство грибов |
Дрожжи и др. |
Нэш [201] |
|
Царство бактерий |
Бифидобактерии, Erwina caratovora, другие бактерии, вызыающие гниль овощей, картофеля и пр., бактериопланктон |
Гончарова [86]; Загорульская [124]; Винберг [58]; 1956 и др. |
|
Царство растений Подцарство низших растений |
Водоросли пресных водоемов |
Липин [165]; Жизнь пресных водоем. СССР [117] |
|
Подцарство высших растений |
Кубышка, картофель, кукуруза, огурцы, ячмень, пшеница, лиственница, береза и др. |
Родченко и др. [242]; Рощин [244]; Нэш [202]; Угаров [304] и др. |
Феномен этот наблюдается на всех уровнях организации живой материи: у органоидов клетки (митохондрии, хлоропласты), клетки, тканей, органов и, наконец, у целых организмов. Анализ приведенных материалов позволяет сделать заключение о том, что резкие изменения интенсивности физиологических процессов, начало которых наблюдается в области 4 °С – это объективно существующее явление, присущее не только растительным, но и всем другим представителям живых организмов. Другими словами, оно имеет общебиологический характер и отражает определенные этапы эволюционного развития живого органического мира Земли. Поскольку активная жизнедеятельность организмов и их переход в состояние вынужденного покоя, оцепенения и спячки, которые ярко прослеживаются у пойкилотермных и гетеротермных живых организмов, тесно связаны с началом теплых и холодных температур, и в связи с этим пограничную температуру – +4 °С можно назвать температурой, где находится биологический нуль.
Таким образом, вся совокупность приведенных нами фактов и теоретических соображений, позволяет констатировать, что состояние живой материи резко меняется при температуре 4 °С: наблюдается прекращение активной жизнедеятельности – питания, роста, размножения; хладнокровные и некоторые теплокровные животные переход в состояние гипобиоза – спячка, оцепенение, диапауза и покой; теплолюбивые растения погибают и др. В связи с этим, есть веское основание считать, что +4 °С является именно той температурой, которую можно назвать биологическим нулем.
При этом следует иметь в виду, что у гетеротермных и пойкилотерных организмов при длительном действии холода, когда температура всего тела достигает температуру 4 °С весь организм реагирует на температуру как одна отдельно взятая клетка. Поэтому у них явление биологического нуля проявляется на уровне целого организма.
Наступление Биологического нуля у целого организма зависит от габаритов объекта и времени действия холода. Дело в том, что у большинства гетеротермных животных вес и размеры тела довольно значительные, поэтому в первую очередь охлаждаются поверхностные ткани и для достижения температуры Биологического нуля на уровне организма требуется определенное время. В природе, в результате длительного закаливающего действия холода, сопряженность работы всех клеток, тканей, даже всех органов у пойкилотермных и гетеротермных организмов, очевидно, встречается довольно часто. Поэтому переход в состояние гипобиоза у них происходит практически безболезненно.
Для гомойотермных организмов о Биологическом нуле речь может идти только относительно отдельной клетки и изолированных тканей, или группы тканей, изолированных от остальной части организма. Как известно, ткань – это совокупность или комплекс клеток, имеющее общее происхождение, сходное строение и выполняющее одинаковую функцию. В связи с таким сходством клеток, определенная ткань у гомойотермных животных может функционировать согласованно как одна клетка, как сказано выше, когда она полностью изолирована от окружающих тканей. Дело в том, что в процессе охлаждения достичь сопряженную работу между всеми клетками, всеми тканями и системами целого организма практически невозможно не только из-за неравномерности охлаждения целого организма, когда поверхностные ткани охладились до температуры биологического нуля, температура глубоко лежащих тканей все еще остается довольно высокой и они будут продолжать активно функционировать, но и из-за их взаимовлияния и регуляторной деятельности нервной и гуморальной систем, закрепленного генетически.
Таким образом, мир живой природы очень разнообразен, однако у них есть одно общее объединяющее начало – это клеточное строение их тела. Согласно клеточной теории строения живых организмов, клетка является элементарной структурно-функциональной и генетической единицей всего живого [210]. В клетке обнаруживаются все важнейшие характеристкики жизни. Другими словами, клетка является наименьшей единицей живого. В связи с этим, вполне логично было предполагать, что Биологический нуль четко проявляется на уровне клеток всех живых организмов.
Теперь попытаемся сформулировать понятие биологического нуля, с учетом всех тех обсуждаемых критериев, которые были перечислены нами выше.
Низкая пороговая температура конца и начала активизации процессов ассимиляции в клетках живых организмов называется биологическим нулем. Такой температурой является +4 °С Цельсия или 0 °U по биологической температуной шкале Угарова.
Примечательно, что температура биологического нуля – +4 °С находится на границе между теплом и холодом, тем самым, температурой инактивации архекомплекса систем жизнедеятельности (АСЖД) и начала активного функционирования системы гипобиометаболизма (СГБМ). Поэтому при температурах ниже биологического нуля гетеротермные и пойкилотермные организмы впадают в состояние гипобиоза, а гомойотермные животные усилиенно генерируют тепло для поддержания постоянства температуры тела, теплолюбивые растительные организмы погибают, вследствие инактивации архекомплекса систем жизнедеятельности и отсутствия системы гипобиометаболизма.
Выше биологического нуля процесс ассимиляции начинает превалировать над процессом диссимиляции, что вызывает активизацию жизнедеятельности и выходу организма из состояния гипобиоза.
Чем же отличается предлагаемое определение биологического нуля?
Во-первых, биологический нуль, по данному определению, относится к клетке, и, поскольку все живые существа состоят из клеток, следовательно, Биологический нуль един для всех живых организмов и находится при температуре 4 °С.
Во-вторых, в качестве биологического процесса, характеризующего состояние жизнедеятельности при биологическом нуле, выбрана активность ассимиляции, которая резко подавляется, а диссимиляция, наоборот, усиливается. Как справедливо считает А.М. Голдовский [82], «неотъемлемыми свойствами (атрибутами) жизнедеятельности должны быть признаны лишь саморегулирующийся двухсторонний обмен (сочетание ассимиляции и диссимиляции) в объеме его основного комплекса и неразрывно связанная с ним реактивность. Именно эти атрибуты характеризуют тот основной процесс, который продолжает протекать в жизнедеятельных состояниях и при выключении всех других проявлений жизнедеятельности – роста, развития, движения, самовоспроизведения. Это, по-видимому, и есть тот основной процесс, познание которого может открыть сущность жизни».
Как было показано выше, при температуре 4 °С, то есть при биологическом нуле, происходит резкое изменение в соотношении между ассимиляцией и диссимиляцией, доходящей до прекращения процесса ассимиляции у организмов. Это приводит гетеротермных и пойкилотермных организмов к гипобиозу, у гомойотермных животных – к усилению термогенерации, а у теплолюбивых растений – к гибели.
Таким образом, новое определение биологического нуля, на наш взгляд, более полно раскрывает его сущность, еще раз доказывая единство всех живых организмов, отражая этапы эволюции органического мира.
Так с усложнением обмена веществ, в связи с адаптацией организмов к холоду и засухе, значение биологического нуля меняется. Например, если на заре начального этапа развития жизни на Земле, биологический нуль (4 °С) можно было охарактеризовать как нижнюю пороговую температуру прекращения жизнедеятельности или температуру гибели организма, вследствие инактивации архесистемы (АСЖД), которая была единственной системой, обеспечивающей всю жзнедеятельность организма.
В настоящее время, когда у части организмов в процессе эволюционного развития появился новый механизм, способный функционировать в среде льдоподобной воды (СГБМ – система гипобиометаболизма), вследствие чего, стало возможным существование пойкилотермных и гетеротермных организмов при температурах ниже 4 °С. Таким образом, эволюционно молодые организмы уже не погибают при понижении температуры ниже +4 °С, однако, начиная с этой температуры у них резко замедляются процессы ассимиляции. Другими словами, у холодо – и морозоустойчивых организмов Биологический нуль находится также при температуре +4 °С. Интересно отметить, что у пойкилотермных и гетеротермных организмов древняя, то есть архесистема жизнедеятельности (АСЖД) сохранялась в неизменном виде и активность процессов ассимиляции в нормальных температурных условиях существования связана именно с ней.
Вторая, эволюционно молодая система гипобиометаболизма (СГБМ) также постепенно совершенствуется и, вполне возможно, появление новых узкоспециализированных организмов, активно живущих в среде льдоподобной воды. Такие организмы, по-видимому, можно будет отнести к организмам, вторично утратившим первый, то есть архекомплекс систем жизнеобеспечения и синтеза (АСЖД), и приспособленным к холодной среде существования. С другой стороны, более вероятным является допущение о появлении у организмов новой системы, разрушающей льдоподобную структуру воды до плотноупакованной. В таком случае отпадает необходимость в создании еще одного нового криокомплекса систем жизнедеятельности, активно функционирующего в условиях холода и мороза. Однако, криофилы в эволюционном плане имеют определенную перспективу, так как на Земле много еще времени и пространства с холодной температурой, где неприспособлены жить большинство нынешних организмов.
И последнее. Если Биологический нуль есть объективно существующее явление, то логично предполагать, что природа должна его активно использовать. Оказалось, что это действительно так.
Особое значение имеет биологический нуль для водных организмов. Давно известно, что при 4 °С вода имеет наибольшую плотность и соответственно наибольшую массу. Когда осенью поверхностный слой воды охлаждается до температуры 4 °С, он опускается на дно озера или реки. А наверху из «легкой» воды, охлажденной до 0 °С, постепенно образуется лед, который прикрывает воду от мороза. При этом на дне глубокого водоема в любой период зимы температура воды будет равна 4 °С, а ближе ко льду – 0 °С. Следует заметить, что даже в условиях многолетней мерзлоты и суровой зимы температура воды в озерах не охлаждается ниже температуры, установленной для озер умеренной широты в зимнее время [93].
В глубоководных озерах слой воды с температурой 4 °С сохраняется летом очень долго и даже постоянно в течение всего года. В пресноводных водоемах обитает значительное количество видов высших растений – такие как рдестовые, наядовые, хвостниковые, сладкоягодниковые и др. Известно также, что виды рясок, вегетировав в течение лета над водой, осенью опускаются на дно водоема и перезимовывают там. Массово произрастающая в водоемах окрестностей г. Якутска ряска малая (Lemna minor L.), по нашим наблюдениям, опускается на дно озера в середине сентября, когда средняя суточная температура воздуха переходит через температуру +4 °С и всплывает на поверхность воды во второй декаде мая. Таким образом, ряска малая благополучно проводит весь неблагоприятный период года под водой, где температура близка к +4 °С или биологическому нулю. Зимой в водоемах Якутии высшие растения и водоросли находятся в состоянии гипобиоза. Наиболее оптимальным температурным условием их переживания является +4 °С, о чем свидетельствует максимальная концентрация зимующего биофонда, наблюдающаяся на больших глубинах, где температура равна к Биологическому нулю и, очевидно, является идеальным местом для переживания организмами неблагоприятных условий среды. Не случайно основная масса планктонных водорослей опускается на дно озер и перезимовывают в комплексе с бентосными [117].
Водные организмы – рыбы и организмы, входящие в состав бентоса, являются пойкилотермными. В результате естественного отбора остались виды, приспособленные к длительной голодовке и гипоксии, а их можно переносить легче всего, очевидно, только в состоянии гипобиоза (оцепенения), когда организм расходует минимум кислорода и энергии, соотвественно и питательных веществ.
По данным Ф.Н. Кириллова [141] в условиях Якутии караси всех возрастных групп зимой не питаются, так как находятся в состоянии оцепенения. Как показывает практика, в такое состояние они впадают в ноябре и начинают активизироваться где-то в апреле-мае. Рыбаки-якуты знают, что зимующего карася надо искать в самом глубоком месте озера, и поэтому такие места летом заблаговременно отмечаются особыми знаками из длинной жерди, чтобы весной точно найти это место и выловить зимующих карасей.
Если температура водоема близка к 4 °С, то можно предположить, что температура тела карася в состоянии оцепенения должна быть близкой также к 4 °С, то есть биологическому нулю. Во время длительного оцепенения, продолжающегося в условиях Якутии не менее 5–6 месяцев, ими мало расходуется энергии, поэтому качество карасей, их питательная ценность остается очень высокой, и якуты до настоящего времени организуют весенний подледный лов карасей. Нерест этих рыб, который требует мобилизации сил и энергии, идет весной почти сразу после схода льда, что также указывает на очень низкий уровень обмена веществ во время оцепенения, в результате которого энергетические ресурсы сохраняются, упитанность существенно не снижается.
Другие обитатели пресноводных водоемов – пауки-серебрянки, как пишет А.С. Константинов [149], в экспериментальных условиях изучения суточного рациона в зимнее время больше всего прожорливы при температуре 27 °С, а при температуре 4 °С пауки уже не питаются. Это, по-видимому, не единичные примеры: о прекращении питания, активной деятельности при температуре, близкой к 4 °С, у водных организмов можно найти в литературе довольно много сведений.
Е.В. Каширская [139] изучала строительную деятельность личинок ручейников. Опыты проводились в чашках Петри, куда помещались личинки ручейников без домиков и различный строительный материал. Определялся промежуток времени, за который личинки, пользуясь предложенным материалом, смогут построить домик для обычных для данного вида размеров. Для выяснения зависимости темпа строительной деятельности от температуры личинкам предлагался наиболее подходящий для особей каждого вида материал при различных температурах (от 0 до 35 °С). Опыты показали, что наиболее эвритермными в этом отношении оказались личинки ручейника двуточечного (Phrygonea bipunctata), продолжавшие сооружение домиков при снижении температуры воды до 4 °С. Личинки Анатолии сильной (Anatolia nervosa) прекращали свою деятельность при температуре 4 °С. У всех видов скорость сооружения домиков была максимальной в интервале температуры от 15 до 25 °С.
Многие земноводные зимуют на дне водоемов [165].
Озерный гольян, как и карась, в зимнее время не питается, а также впадает в спячку. По наблюдениям рыболовов, в условиях Якутии такое оцепенение у гольянов начинается раньше, чем у карасей – в октябре-ноябре. При этом интересно, что эта рыба зимует не поодиночке, а собирается в довольно большую группу, которую рыбаки называют «стаей» и опускается на дно водоема, где и проводит зиму. Температура тела у них, по-видимому, приближается к биологическому нулю, во всяком случае, расход энергии, как и у карасей, ничтожен, о чем свидетельствует высокая упитанность гольянов к концу зимовки, что позволяет якутам весной в марте-апреле организовать их подледный лов специальным сачком-куйууром.
Обязательные обитатели пресных водоемов – амебы при 4 °С не способны делиться: ни цито- ни кариокинез не происходят, а синтез ДНК протекает очень медленно [384].
А.М. Рощиным [244] изучался окислительный метаболизм корневищ кубышки желтой (Nuphar Luteum) при воздействии на них различными факторами. Автором обнаружено постепенное увеличение интенсивности дыхания до 2–5 раз по сравнению с первоначальной – при инкубации тонких срезов корневищ кубышки при комнатной температуре.
Такую картину раньше наблюдали исследователи на срезах подземных запасающих органов растений, произрастающих на суше. Было установлено, что многие факторы, нарушающие нормальный метаболизм срезов запасающих органов, предотвращают усиление дыхания во время инкубации. К ним относятся, как пишет автор, ингибиторы металлосодержащих оксидаз (цианид, азид, окись углерода), при нормальной температуре ионы лития, конкурентно замещающие ионы калия, а также хлорамфеникол, являющийся ингибитором синтеза белка.
Интересно отметить, что к числу подобных мощных факторов, нарушающих нормальный метаболизм срезов, относится также понижение температуры в интервале 0–4 °С. Проведенные А.М. Рощиным исследования по изучению влияния температуры 3 °С на развитие инкремента дыхания показали, что пониженная температура предотвратила увеличение интенсивности дыхания, в то же время поглощение кислорода срезами при комнатной температуре за 6 часов возросло в 3 раза. В присутствии цианида калия (5∙10–3 М) интенсивность дыхания также не увеличивается.
Примеров о влиянии пониженной температуры от 1 до 4 °С на жизнедеятельность простых водных организмов можно было бы привести много, но даже сказанное достаточно ярко иллюстрирует неблагоприятность этой зоны температуры для активной жизни. О влиянии постоянной температуры 4 °С водоема на жизнедеятельность водных организмов можно судить по содержанию растворенного кислорода, поскольку образование кислорода и его окисление, если не учитывать известных условий растворения кислорода в воде, в основном зависит от деятельности растительных и животных организмов, заселяющих данный водоем.
В озере субальпийского типа, примерно, начиная с 50 м, температура воды постоянна и равняется 4 °С. Кривая содержания кислорода в воде, начиная с 50-метровой глубины, практически не меняется. Это, очевидно, указывает на низкий уровень образования и окисления кислорода, то есть все биологические процессы, идущие с поглощением и выделением кислорода, в указанных условиях резко снижены и область водоема с такой температурой заселена живыми организмами слабо.
Таким образом, можно предположить, что зона пресноводного водоема с температурой 4 °С является зоной «дремлющей» жизни. Разумеется, эту зону называть «мертвой» нельзя, так как при температуре 4 °С могут прекрасно себя чувствовать как эвритемные виды, сохраняющие активность даже при более низких температурах, чем эта, так и виды, впадающие в спячку или в оцепенение (караси, гольяны и др.). Дело в том, что спячка или оцепенение не являются простыми процессами, зависящими только от температурного фактора. Это необходимый сезонноповторяющийся биологический процесс или биоритм, обусловленный главным образом, внутренним механизмом, геномом, для срабатывания которого температура среды является внешним пусковым сигналом. Например, в осеннее время водные организмы, впадающие в оцепенение, довольно длительное время могут находиться в активном состоянии при температуре, близкой к 4 °С. Правда, они могут выбирать в это время и другую температуру, градиент которой от кромки льда до дна водоема составляет от 0 до 4 °С.
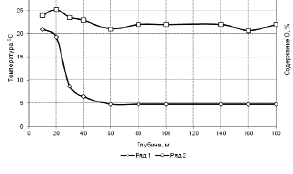
Рис. 8.2.1. Вертикальное распределение температуры (2) и кислород, (в процентах от состояния насыщения) (1) в озере Сенека субальпийского типа [165]
Как пишут В.А. Аминева, А.А. Яржомбек [13] лещ, синец, плотва и другие обитатели наших рек летом стремятся к температуре 27 °С, а зимой – к температуре 2–3 °С, при которой лучше происходит зимовка.
Представляет интерес данные, полученные у амеб при различных контролируемых температурных условиях. При содержании культур амеб (Amoebae proteus) в условиях 1–4 °С за 15 суток делилось всего лишь 2 амебы из 100, при 7–11 °С через 4 дня разделились 70 особей из 100, а на 5-й день – 88 особей. Интересно отметить, что у амебы (Amoebae proteus) вязкость цитоплазмы при температуре 4,5 °С максимальна [280].
Естественно думать, что зимоспящие животные должны поддерживать температуру своего тела на уровне биологического нуля. Анализ литературных данных показывает, что это, действительно, так. Например, если суслики в активном состоянии имеют температуру тела, равную 36,8–38,0 °С, то во время сна температура тела зверьков понижается до 3,6–4,8 °С [31]. Как пишут Н.Г. Соломонов и др. [267] длиннохвостый суслик, по показаниям термистора, вживленного в межлопаточную область, впадает в спячку при понижении температуры тела ниже 5 °С.
Более подробно остановимся на данных Т. Серкиной и В. Поповой (работа проводилась под руководством к.б.н. М.К. Слепцова и к.б.н. А.К. Ахременко), полученных в экспериментальных условиях в течение зимы у 72 экземпляров длиннохвостого суслика (Citellus undulatus). Температура регистрировалась ректально при помощи медицинского термометра и медь-константовой термопары. Температура помещения (подвала), где содержались зверьки, и где проводился эксперимент, была близкой к 2 °С [258].
Ректальная температура спящих животных в начале декабря составила 13,5 ± 0,43 °С, а в период глубокого сна снижалась до 3,75 ± 0,44 °С и на таком уровне держалась на протяжении января и февраля (рис. 8.2). В апреле температура тела сусликов поднялась до 31,8 ± 0,42 °С, и они скоро пробудились.
Самым интересным здесь является то, что во время настоящей спячки температура тела у суслика поддерживается на уровне биологического нуля, хотя температура окружения примерно на 2 °С ниже. Возникает вопрос, почему температура тела у суслика поддерживается именно на уровне биологического нуля и как? На первый вопрос мы уже ответили: при температуре биологического нуля расход энергии на поддерживание жизни минимален, чем при всех других температурах. Это имеет немаловажное значение для переживания организмом неблагоприятного периода года, который длится в умеренных и особенно северных широтах довольно долго, например, в Якутии до 7–8 месяцев. Теперь постараемся ответить на второй вопрос. Когда под действием холода температура тела спящего суслика опускается ниже 4 °С, включается известная нам вторая система жизнеобеспечения – (СГБМ). При этом повышаются обменные процессы, интенсивность дыхания и температура тела зверька соответственно поднимается. Однако по достижении температуры тела до 4 °С интенсивность деятельности второй системы жизнеобеспечения сильно падает, а холодный воздух и холодные тела, окружающие животное, снова снижают температуру его тела. Когда температура тела животного вновь падает ниже биологического нуля, снова включается вторая система жизнеобеспечения, в результате чего, температура тела снова повышается и так повторяется многократно весь период спячки суслика. Таким образом, у организма автоматически поддерживается температура биологического нуля по принципу работы термостата, где биологический контактный термометр установлен на отметке 4 °С, то есть на биологическом нуле. Сказанное проиллюстрируем экспериментальными данными, почерпнутыми из литературных источников.
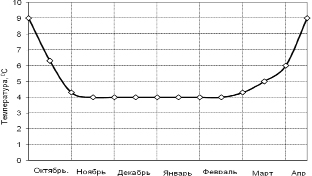
Рис. 8.2.2. Динамика ректальной температуры тела суслика в период зимней спячки [258]
Н.И. Калабухов [135] приводит данные опытов Кайзера по дыханию сусликов при различных температурах. В этих опытах суслики в спячке при 5 °С в течение часа потребляли на 1 кг массы 19,7 см3 кислорода, при понижении температуры воздуха до 2 °С – почти вдвое больше – до 38,9 см3. Это, естественно, приводило к пробуждению животных.
Н.Г. Соломонов и др. [267] отмечают, что в процессе охлаждения, когда температура тела суслика в межлопаточной области падает ниже 4 °С, животное пробуждается. М.Б. Штарк [337] в своей книге «Мозг зимоспящих» пишет: «Нам не удавалось наблюдать снижения температуры более чем до 2 °С для корки головного мозга и 3 °С – для корковых отделов. Более значительное падение внутримозговой температуры сопровождалось полным или «частичным» пробуждением животного» (с. 60). Естественно, пробуждение не состоялось бы без усиления обменных процессов и повышения температуры мозга и тела животных.
В опытах Е.В. Майстраха [168] крыс охлаждали в 2 стадии. Вначале нормальных крыс сажали в герметически закрытые сосуды, а сосуды помещали в холодильник с температурой около 5 °С на 2–2,5 часа (первая стадия). Затем животные извлекались из сосудов на свежий воздух и быстро переносились в кашицу из мелко наколотого льда, где и охлаждались до температуры тела, приближающейся к 0 °С (вторая стадия).
В этих экспериментах интересно то, что ректальная температура во второй стадии охлаждения снижается неравномерно. Так, на протяжении первых 5–10 мин, как пишет автор, она падает медленно. Начиная с 19 °С, ректальная температура снижается экспоненциально времени охлаждения, и после достижения 5 °С падение ректальной температуры вновь замедляется. Другими словами, наблюдается некоторое сопротивление дальнейшему опасному для жизни падению температуры, очевидно, путем выработки тепла. У крысы, как у незимоспящего животного, система гипобиометаболизма (CГПБМ), разумеется, не совершенна, поэтому животное не пробуждается и гибнет от холода. Легко понять, что если бы не было механизма термогенеза, точнее второй системы жизнеобеспечения у зимоспящих животных, то они не пробудились бы при охлаждении их тела до опасных для жизни низких температур и погибли бы как эта крыса от замерзания. По-видимому, наиболее чувствительная к падению температуры ниже 0 °С регуляторная ткань у зимоспящих находится в их мозгу.
Повышение потребления кислорода при падении температуры ниже 4 °С наблюдается и у водных организмов. Так в январе интенсивность процессов метаболизма у моллюска литторина (Littorina littorea) минимальна при 4 °С, а при охлаждении тела примерно до 1 °С повышается до уровня, который может наблюдаться при температуре 15 °С [379] Можно полагать, что литторина в зимнее время также поддерживает температуру своего тела на уровне биологического нуля.
По-видимому, биологический нуль, где это возможно, широко используется пойкилотермными животными в период переживания неблагоприятного холодного сезона года.
Термогенез или выработка тепла происходит у организмов в результате повышения уровня обмена веществ, в частности, окислительных процессов дыхания. Поэтому усиление выделения тепла при понижении температуры ниже 4 °С наблюдается и у растительных организмов. Разумеется, термогенез у растений не имеет такого большого значения для их жизни, как у животных, однако может играть определенную роль во время кратковременных заморозков, не давая опускаться температуре тканей до опасных повреждающих величин. В этом отношении интересны наблюдения А.К. Винтер [59], которая установила, что 5-часовое действие искусственного ночного заморозка с минимальной температурой до –4 °С не снижало температуру листьев кукурузы до отрицательных значений. Этот факт не может быть объяснен только тепловой инерцией, вызванной массивностью растения, хотя вклад его и нельзя отрицать. Главной причиной относительной стабильности температуры надземной части растения в данном случае во время заморозков является, скорее всего, термогенез.
Как пишут Г.Ш. Нахуцришвили, З.Г. Гамцемлидзе [198], при небольших отрицательных температурах воздуха температура растения почти всегда остается положительной, при этом температура стеблей и цветков ниже температуры листьев.
Наши наблюдения за температурой цветка весеннего эфемероида – прострела желтеющего, массово произрастающего в Центральной Якутии, показало, что ночью температура цветка падает параллельно с падением температуры окружающего воздуха, однако, начиная с 4 °С эта картина нарушается температура воздуха падает быстрее, чем температура цветка [315].
После всего вышесказанного, является вполне естественным и закономерным устройство зимней гнездовой камеры зимоспящими на такой глубине, где температура равна или близка к биологическому нулю. Например, Д.И. Бибиков [29] сообщает, что «зимние гнездовые камеры в межгорных выровненных долинах высокогорного Тянь-Шаня и Памиро-Алая сурки устраивают на глубине 2 м и более, где температура наиболее близка к благоприятной для спячки (5 °С). В высокогорье Алая она располагается на глубине 3,6 м, а у нижнего высотного предела распространения красного сурка – 1,8 м» (с. 283). Таким образом, где это возможно, то есть в тех местах, где отсутствует многолетняя мерзлота, зимоспящие для своей спячки ищут своеобразное «дно водоема». Но в отличие от водных организмов у них имеется совершенная система терморегуляции, работающая во время спячки, поэтому они сумели освоить даже высокие широты, где царствует многолетняя мерзлота и длинная, суровая зима. На севере Якутии, где из-за вечной мерзлоты зимняя камера располагается неглубоко, сурки утолщают ее потолок, штукатурят стенки, применяют дополнительные травяные пробки для лучшей теплоизоляции. Несмотря на это, к февралю-марту температура в камере падает до –14 – 22 °С, что приводит к частому пробуждению зверьков, усиленному расходу ими жира и увеличению смертности [30, 18, 267].
Периодические пробуждения суслика с повышением температуры тела до температуры бодрствующего состояния были зафиксированы при помощи телеметрической установки [400]. Термодатчики были установлены в норах 12 канадских сусликов (Citellus richardsoni) и температура регистрировалась через каждые 10 мин. с июля по март. Было установлено, что длительность периодического прерывания спячки с осени до января возрастает, а затем снова убывает. Раньше такие же данные были получены и другими исследователями в лабораторных условиях [384, 357]. Это, очевидно, результат работы «биологического термостата», в прямом смысле, спасющего животных от замерзания, то есть от их верной гибели во время спячки.
Таким образом, Биологический нуль активно «эксплуатируется» природой.
Следует также отметить, что температура 4 °С еще может быть принята за биологический нуль чисто условно. Как известно, при создании шкалы Цельсия в качестве условной нулевой точки отсчета принята температура замерзания воды, а температура ее кипения условно принята за 100 °С. При этом, несомненно, очень большую роль сыграло то обстоятельство, что вода является очень распространенным минералом в природе. А живой организм на 80–90 % состоит из воды. Считается, что вода является матрицей жизни, участницей и средой фундаментальных биохимических реакций. Поэтому поистине замечательная аномальная точка воды – температура наибольшей плотности, то есть 4 °С, при которой происходит своего рода фазовый переход – состояние воды резко меняется, и она превращается в жидкий лед, играющий огромное значение в жизнедеятельности организмов, вполне может быть принята за биологический нуль, как сказано выше, даже чисто условно.
Все сказанное выше в совокупности делает, как нам кажется, предлагаемую концепцию о биологическом нуле, едином для всего живого мира Земли, вполне убедительной.