
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

ПАРАТГОРМОН-РОДСТВЕННЫЙ ПРОТЕИН. 2-е издание переработанное и дополненное
Курзанов А. Н., Ледванов М. Ю., Быков И. М., Медведев В. Л., Стрыгина Е. А., Бизенкова М. Н., Заболотских Н. В., Ковалев Д. В., Стукова Н. Ю.,
14.2.2. ПТГрП-сопряженные сигнальные механизмы морфогнеза молочных желез
Молекулярные события, лежащие в основе раннего развития молочных желез, определяющие эти важнейшие эпителиально-мезенхимных взаимодействия осуществляются при участии ПТГрП, фактора роста фибробластов, эпидермального фактора роста, сигнальных путей IHH, SHH, Wnt/β-катенин и играют важную роль в формировании исходных плакод молочных желез. (Chu E.Y. et al., 2004; Davenport T.G. et al., 2003; Hatsell S.J., Cowin P., 2006; Veltmaat J.M. et al., 2006). Передача сигналов ПТГрП, по-видимому, имеет решающее значение для спецификации мезенхимы молочной железы, которая, в свою очередь, поддерживает эпителиальную идентичность молочной железы, направляет рост почек, нарушает рудимент мужской молочной железы и определяет образование соска (Hiremath M., Wysolmerski J., 2013). Сигнальный путь Hedgehog был первоначально обнаружен как критический путь для формирования паттерна и развития эмбриона (Wu F. et al., 2017). Несколько исследований ранее идентифицировали передачу сигналов hedgehog в качестве активатора активации ПТГрП в развитии кости и хряща, а также рака молочной железы (Jemtland R. et al., 2003; Sterling J.A. et al., 2006; Katoh Y. and Katoh M., 2009). Кроме того, передача сигналов Hedgehog, как было ранее показано, важна для морфогенеза молочной железы во время эмбрионального развития молочной железы, а также в начале полового созревания (Moraes R.C. et al., 2007; Okolowsky N. et al., 2014). Продемонстрировано, что ПТГрП регулирует дифференцировку и функцию мезенхимы молочной железы, частично, посредством активации передачи сигналов Wnt специфически в мезенхимальных клетках. Доказано, что каноническая передача сигналов Wnt необходима для нормального развития эмбриональных молочных желез, для образования плакод молочных желез и развития эмбриональной системы молочных протоков. Показано, что множественные Wnt-сигнальные пути, Wnt-рецепторы и Wnt-модуляторы/ингибиторы экспрессируются эпителиальными и/или мезенхимными клетками зачатка молочной железы (Chu E.Y. et al., 2004; Boras-Granic K. et al., 2006).
Факторы, участвующие в этом сигнальном пути, такие как Wnt10b, Lef1 и β-катенин, экспрессируются в развивающизся молочных железах (Chu E.Y. et al., 2004; Veltmaat J.M. et al., 2004). Наконец, было показано, что генетические изменения в каноническом сигнальном пути Wnt нарушают нормальное эмбриональное развитие молочных желез. Сверхэкспрессия Wnt-ингибитора Dkk1 в эмбриональном эпидермисе Трансгенных мышей in vivo полностью блокирует образование плакод молочной железы. Эти данные указывают на то, что передача сигналов WNT способствует развитию плакод и необходима для инициации морфогенеза молочной железы (Chu E.Y. et al., 2004). Передача сигналов ПТГрП индуцирует экспрессию Lef1 и β-catenin в мезенхиме молочной железы, что позволяет предположить, что ПТГрП может модулировать каноническую передачу сигналов Wnt в этих клетках (Dunbar M.E. et al., 1999). Показано, что каноническая передача сигналов Wnt/ β-catenin активируется в конденсирующейся мезенхиме во время формирования зародышевых почек молочных желез (Hiremath M. et al., 2012). Передача сигналов мезенхимального Wnt требует действия ПТГрП и β-катенина, но не Lef1, и способствует правильной пролиферации, дифференцировке и функции мезенхимы молочной железы. Удаление мезенхимальных β-катениновых фенокопий является критическим элементом потери ПТГрП и PTH1R и блокирует эффекты избыточной экспрессии ПТГрП.
Доказано, что передача сигналов Wnt активируется в конденсирующейся мезенхиме в ответ на ПТГрП, который секретируется из эпителия. Кроме того, активация передачи сигналов Wnt в мезенхимальных клетках необходима для того, чтобы они полностью дифференцировались в ответ на передачу сигналов ПТГрП. Потеря β-катенина только в мезенхиме молочной железы приводит к образованию молочной почки с меньшим количеством окружающих мезенхимальных клеток. По-видимому, это является результатом снижения пролиферации мезенхимальных клеток, а не увеличения скорости апоптоза. Кроме того, потеря β-катенина ингибирует экспрессию рецептров эстрогенов и андрогенов и Lef1 которые обычно отвечают на передачу сигналов ПТГрП (Hiremath M. et al., 2012). Потеря мезенхимального β-катенина влияла на способность мезенхимы поддерживать андроген-опосредованное разрушение молочных почек у мужских эмбрионов. Утверждается, что передача сигналов Wnt действует ниже ПТГрП, стимулируя пролиферацию, дифференцировку и функцию мезенхимальных клеток. Молекулярные механизмы, с помощью которых ПТГрП активирует передачу сигналов β-катенина в конденсирующейся мезенхиме молочной железы, остаются неясными, но могут включать аутокринную или паракринную активацию канонического пути Wnt. Эти результаты демонстрируют активацию как белка R-спондина 1, так и Wnt11 в ответ на воздействие ПТГрП. Предыдущие исследования показали, что оба этих гена специфически экспрессируются мезенхимными клетками в зародышах молочной железы эмбрионов (Chu E.Y. et al., 2004; Nam J.S. et al., 2007). Белки R-спондина усиливают каноническую передачу сигналов Wnt, взаимодействуя с высокоаффинным трансмембранным рецептором Dickkopf 1 (DKK1), который функционально блокирует передачу сигналов WNT/beta-catenin. R-спондин является компонентом мембранного комплекса, который модулирует каноническую передачу сигналов WNT через белок, связанный с рецептором липопротеина LRP6, позволяя связываться с Wnts и Frizzleds (Binnerts M.E. et al., 2007; Kim K.A. et al., 2008). Хотя Wnt11 часто считают «неканоническим» лигандом Wnt, он также может активировать канонический путь. В остеобластах ПТГрП усиливает экспрессию R-спондина 2, что облегчает каноническую передачу сигналов Wnt с помощью Wnt11 (Friedman M.S. et al., 2009). Следовательно, возможно, что ПТГрП активирует передачу сигналов Wnt в зародыше молочной железы посредством активации Wnt11 и белка R-спондина 1 в мезенхиме молочной железы. Кроме того, активация передачи сигналов Wnt в мезенхиме молочной железы с помощью ПТГрП возможно включает через белок, связанный с рецептором липопротеина LRP6. Таким образом, установлено, что передача сигналов мезенхимального β-катенина имеет решающее значение для нормального развития зародыша молочной железы и для экспрессии ключевых молекулярных маркеров мезенхимы молочной железы. Кроме того, активация передачи сигналов Wnt/β-catenin в этих клетках происходит ниже пути передачи сигналов ПТГрП/PTH1R.
Hens J.R. et al., (2007) изучили взаимодействие между сигналами ПТГрП и BMP во время развития ранних эмбриональных молочных желез. Авторы продемонстрировали, что сигнализация ПТГрП является разрешительной для сигнализации BMP в мезенхиме молочной железы. Показано, что ПТГрП действует, по крайней мере, частично, путем модуляции передачи сигналов BMP в клетках мезенхимы молочной железы. При образовании почек молочной железы BMP4 экспрессируется мезенхимными клетками на вентральной поверхности эмбриона. Передача сигналов ПТГрП сенсибилизирует мезенхимальные клетки к BMP in vitro и in vivo, увеличивая экспрессию рецептора BMP1A. Взаимодействие между ПТГрП и BMP4 важно для прорастания почки молочной железы. Комбинация сигналов ПТГрП и BMP увеличивает экспрессию гомеобоксного гена MSX2 в мезенхиме молочной железы, что, в свою очередь, позволяет ПТГрП подавлять образование волосяных фолликулов в зоне развивающейся почки молочной железы и соска. Полученные данные свидетельствуют, что сигнальная передача ПТГрП активирует экспрессию BMPR1A в мезенхиме молочной железы. Это взаимодействие, в свою очередь, активирует экспрессию гена Msx2 в мезенхимальных клетках, что позволяет им подавлять образование волосяных фолликулов в коже, покрывающей соски. ПТГрП регулирует в мезенхиме молочной железы экспрессию MSX2, который является транскрипционным фактором, участвующим в эпителиально-мезенхимальных взаимодействиях во время развития. Считается, что он имеет важные функции при формировании эпидермальных придатков, поскольку нарушение гена Msx2 приводит к аномальному развитию зуба, волосяного фолликула и молочной железы (Satokata I. et al., 2000). Эти данные определяют MSX2 как важный мезенхимальный фактор, который позволяет ПТГрП и BMP4 подавлять индукцию волосяного фолликула в локусе развивающейся почки молочной железы и соска. Сигналы ПТГрП и BMP также взаимодействуют, чтобы мезенхима могла инициировать рост эпителия почек молочной железы. Присутствие специфического BMP необходимо для полного осуществления эффектов ПТГрП. В случае нормального развития молочной железы экспрессия BMP4 в вентральной мезенхиме совпадает с образованием почек молочной железы, которые экспрессируют ПТГрП и позволяют взаимодействовать с сигнальными путями для образования мезенхимы, специфичной для молочных желез. Результаты исследования Hens J.R. et al., (2007) показывают, что ПТГрП-опосредованная экспрессии BMPR1A обеспечивает пространственно ограниченную, аутокринную или паракринную сигнализацию BMP внутри мезенхимы, позволяющую клеткам мезенхимы молочной железы запускать и/или поддерживать рост эпителия зародышевой почки молочной железы. В дополнение к стимулированию роста почек, мезенхима, специфичная для молочных желез, индуцирует формирование из эпидермиса кожи сосков, что также зависит от передачи сигналов ПТГрП. Характерной особенностью кожной оболочки сосков является отсутствие у нее волос, что, вероятно, является результатом торможения образования волосяных фолликулов ПТГрП, выделяемого эпителиальной почкой. В отсутствие ПТГрП или PTH1R волосяные фолликулы развиваются слишком близко к почке молочной железы, а в присутствии ПТГрП развитие волосяного фолликула подавляется по всему вентральному эпидермису (Foley J. et al., 2001; Wysolmerski J.J. et al., 1994). Способность ПТГрП подавлять развитие волосяного фолликула зависит от действий фактора транскрипции гомеобокс MSX2. Используя гибридизацию in situ, авторы обнаружили, что MSX2 экспрессируется в мезенхиме молочной железы (Phippard D.J. et al., 1996; Satokata I. et al., 2000). Кроме того, мезенхимальная экспрессия MSX2 требует передачи сигналов ПТГрП. Как и в молочной железе, индукция волосяного фолликула требует взаимодействий между эпителиальными и мезенхимальными клетками. Hens J.R. et al. (2007) также обнаружили, что ПТГрП и BMP4 взаимодействуют для подавления развития волос путем индуцирования Msx2 в мезенхиме молочной железы. Результаты экспериментов, представленные авторами, демонстрируют важное взаимодействие между ПТГрП и BMP4 во время развития эмбриональной почки молочной железы (рис. 18).
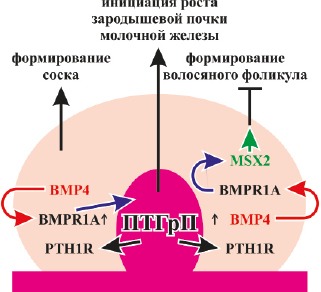
Рис. 18. Взаимодействие паратгормон-родственного протеина и костных морфогенетических белков (ВМР) в инициации роста зародышевой почки молочной железы и формировании соска во время эмбрионального развития
ПТГрП секретируется из эпителиальных клеток молочных желез и повышает экспрессию рецептора ВМР (BMPR1А) в мезенхиме молочной железы. Это повышает чувствительность этих клеток к ВМР и позволяет им реагировать на BMP4 паракринно и/или аутокринно Передача сигналов ПТГрП через PTHR1 также активирует BMPR1А и сенсибилизирует мезенхимальные клетки молочной железы к BMP4, возможно, усиливая передачу сигналов BMP в мезенхиме молочной железы. Этот путь необходим для инициации роста зародышевой почки молочной железы через MMP2 и для подавления формирования волосяного фолликула в соске, индуцируя экспрессию фактора транскрипции гомеобокс Msx2, что приводит к ингибированию образования волосяных фолликулов в развивающейся оболочке соска. Передача сигналов ПТГрП также поддерживает эпителиальную идентичность молочной железы посредством передачи сигналов в мезенхиме молочной железы. В эмбриональной почке молочной железы ПТГрП оказывает влияние на морфогенез, частично, через эпителиально-стромальные перекрестные взаимодействия, опосредуемые сигналами Wnt и BMP.