
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

4.4. Малые деревья ели на пробной площади квартала 30
В табл. 4.3 представлены результаты измерений численности подроста ели, то есть малых деревьев ели на пробной площади квартала 30.
Таблица 4.3
Малые деревья ели , шт.
|
Расстояние от центра |
Геодезические направления |
Всего |
|||||||
|
С |
С-В |
В |
Ю-В |
Ю |
Ю-З |
З |
С-З |
||
|
0 |
9 |
9 |
9 |
9 |
9 |
9 |
9 |
9 |
9 |
|
10 |
13 |
3 |
13 |
2 |
4 |
1 |
1 |
2 |
39 |
|
20 |
15 |
0 |
0 |
14 |
18 |
8 |
10 |
13 |
78 |
|
30 |
9 |
0 |
0 |
6 |
5 |
18 |
14 |
12 |
64 |
|
40 |
17 |
4 |
2 |
12 |
7 |
14 |
8 |
30 |
94 |
|
50 |
15 |
11 |
7 |
7 |
0 |
45 |
20 |
34 |
139 |
|
60 |
12 |
41 |
15 |
55 |
20 |
30 |
11 |
33 |
217 |
|
70 |
5 |
26 |
8 |
49 |
15 |
33 |
10 |
16 |
162 |
|
Итого |
95 |
94 |
54 |
154 |
78 |
158 |
83 |
149 |
802 |
На каждом геодезическом направлении общая закономерность не изменилась. Различия наблюдаются только в соотношениях значений параметров модели. Этот факт указывает на единообразие в поведении ели любого возраста. Ель любого возраста растет организованно.
Север. После структурно-параметрической идентификации была получена трехчастная биотехническая закономерность (рис. 4.20) вида
![]()
![]()
![]() . (4.19)
. (4.19)
|
Рис. 4.20. График изменения количества малых деревьев ели в зависимости от расстояния в северном направлении |
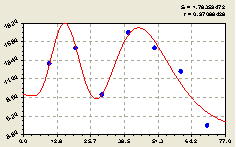
Первая оставляющая формулы (4.19) показывает общую тенденцию экспоненциального спада численности у малых деревьев ели в зависимости от расстояния L
Вторая составляющая характеризует влияние материнской ели на распределение молодого подроста.
По графику на рис. 4.20 видно, что на расстоянии 30 м находится вторая взрослая ель. Поэтому изменение численности малых деревьев ели по третьей составляющей будет проходить, по всей видимости, под влиянием второй взрослой ели. Но при этом заметно, что вторая ель как бы мешает первой, снижая кризисно (отрицательный знак перед третьей составляющей моделиризисно ()метно, что вторая ель как бы мешает первой, снижая ьник.жную конструкцию во многих геодезических направлениях. гляка) численность молодняка.
Северо-восток. По геодезическому направлению снова была получена трехчастная биотехническая закономерность (рис. 4.21) вида
![]()
![]()
![]() . (4.20)
. (4.20)
|
Рис. 4.21. График изменения количества малых деревьев ели в зависимости от расстояния в северо-восточном направлении |
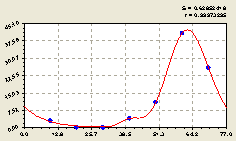
На рис. 4.21 видно, что на расстоянии 50...70 м от материнской ели происходит резкий рост численности малых деревьев ели.
Близко к центральной ели происходит колебательное возмущение с большой амплитудой.
Причем до всплеска численности подроста ели происходит сложное изменение на расстоянии до 50 м от центра.
Восток. После проведения структурно-параметрической идентификации получили трехчастную биотехническую закономерность (рис. 4.22) известного из предыдущих примеров вида
![]()
![]()
![]() . (4.21)
. (4.21)
|
Рис. 4.22. График изменения количества малых деревьев ели в зависимости от расстояния в восточном направлении |
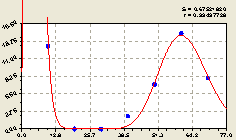
Первая составляющая формулы (4.21) показывает общую тенденцию экспоненциальной гибели малых деревьев ели относительно центрального материнского дерева в зависимости от расстояния L Однако малые деревца испытывают значительные трудности, на что указывает резкие изменения графика на рис. 4.22.
Очевидно, тенденция экспоненциальной гибели молодого подроста, растущего в непосредственной близости от материнской ели, связана с тем, что крупная ель препятствует росту и развитию молодого подроста. Это проявляется в недостатке света, тепла, питания и др. Поэтому, по-видимому, существует оптимальный интервал репродукционного возраста у материнской ели.
Из графика на рис. 4. 22 видно, что по мере удаления от центральной материнской ели численность малых деревьев ели возрастает.
Юго-восток. После структурно-параметрической идентификации была получена биотехническая закономерность (рис. 4.23) вида
![]()
![]() . (4.22)
. (4.22)
|
Рис. 4.23. График изменения количества малых деревьев ели в зависимости от расстояния в юго-восточном направлении |
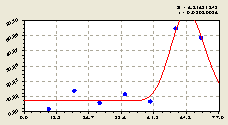
Из графика на рис. 4.23 видно, что в данном геодезическом направлении в непосредственной близости с материнской елью происходит колебательное возмущение с большой амплитудой.
Однако почти до 60 м от материнской ели численность подроста мала. Может оказаться, что здесь должна была влиять другая материнская ель, растущая не более чем на 80-100 м от рассматриваемого дерева. Тогда можно предположить, что в качественном ельнике материнские ели образуют своеобразную сетку из производителей потомства.
Репродуктивная лесная сетка, конечно же, образуется в зависимости от направления преобладающих ветров.
Между тем на расстоянии 50-70 м происходит скачок численности малых деревьев ели. Это, вероятнее всего, объясняется тем, что взрослая материнская ель на этом расстоянии уже не оказывает влияния на рост и развитие молодого подроста.
Можно предположить, что свое влияние здесь оказывает направление господствующего ветра и освещенность территории. Это будет являться целью дальнейших исследований.
Юг. В этом направлении была получена трехчастная модель (рис. 4.24) в виде биотехнической закономерности
![]()
![]()
![]() . (4.23)
. (4.23)
По первой составляющей формулы (4.23) происходит снижение численности малых деревьев ели относительно центрального дерева в зависимости от расстояния L
|
Рис. 4.24. График изменения количества малых деревьев ели в зависимости от расстояния в южном направлении |
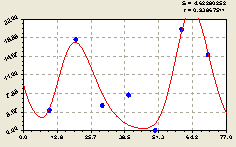
Рядом с материнской елью происходит колебание численности малых деревьев ели с незначительной амплитудой.
В целом, молодой подрост относительно материнской ели в данном направлении расположен неравномерно.
Юго-запад. Поэтому направлению получили биотехническую закономерность (рис. 4.25) вида
![]()
![]()
![]() . (4.24)
. (4.24)
|
Рис. 4.25. График изменения количества малых деревьев ели в зависимости от расстояния в юго-западном направлении |
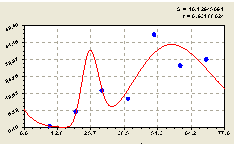
Как и в южном направлении, по первой составляющей формулы (4.24) происходит снижение численности малых деревьев ели.
На расстоянии 40-70 м от материнской ели происходит возмущение численности молодого подроста с большой амплитудой.
Запад. После проведения структурно-параметрической идентификации снова была получена трехчастная биотехническая закономерность (рис. 4.26) вида
![]()
![]()
![]() . (4.25)
. (4.25)
Как и в южном направлении, первая составляющая формулы (4.25) показывает снижение численности молодого подроста, растущего вблизи материнской ели.
На расстоянии 40 м наблюдается рост численности малых елей. Здесь происходит их колебательное возмущение с большой амплитудой.
Северо-запад. Здесь получили формулу (рис. 4.27) вида
![]()
![]()
![]() . (4.26)
. (4.26)
|
Рис. 4.26. График изменения количества малых деревьев ели в зависимости от расстояния в западном направлении |
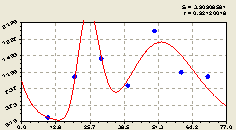
По рис. 4.27 видно, что численность молодого подроста в данном геодезическом направлении изменяется неравномерно. На такое распределение, в первую очередь, оказывает влияние направление ветра и освещенность территории. Но почему на краю круговой пробной площади резко снижается численность подроста ели? Может оказаться, что здесь произошло антропогенное, а может даже техногенное не только от людей, но и от лесных зверей, влияние на возобновляемость ели и её распределение по территории круговой площади на северо-запад.
|
Рис. 4.27. График изменения количества малых деревьев ели в зависимости от расстояния в северо-западном направлении |
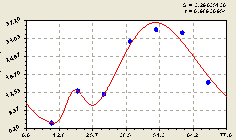
Первая составляющая формулы (4.26) характеризует снижение числа молодых елей, растущих рядом с материнской елью.
На расстоянии 40...60 м происходит значительное колебание молодого подроста с большой амплитудой.
Всего на концентрических кольцах. По концентрическим кольцам круговой пробной площади численность деревьев ели по формуле
![]() (4.27)
(4.27)
распределилась снова, как и всех деревьев и отдельно деревьев вида ели, по закону экспоненциального роста.
|
Рис. 4.28. График изменения количества малых деревьев ели в зависимости от расстояния |
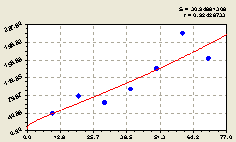
Взрослая ель становится здесь доминирующим растением на круге радиусом, по крайней мере, не менее чем на 60 м распределяя свое молодое потомство.
Все остальные деревья подчинены этой центральной ели. Это доказывается волнообразным расположением точек, то есть подроста, на рис. 4.28.