
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

4.6. Все деревья на пробных площадках
Север. В ходе проведения структурно-параметрической идентификации по данному геодезическому направлению была получена трехчастная биотехническая закономерность (рис. 4.30) вида
![]()
![]()
![]() (4.28)
(4.28)
По первой составляющей формулы (4.28) наблюдается снижение численности всех деревьев относительно материнской ели в зависимости от расстояния L
Вторая составляющая характеризует влияние материнской ели в виде дерева-лидера на рост и развитие ели и других видов деревьев.
На расстоянии 40 м от центрального материнского дерева находится вторая взрослая ель. Поэтому, очевидно, стрессовое возбуждение по третьей составляющей проходит уже под влиянием второй взрослой ели.
|
Рис. 4.30. График изменения количества деревьев на пробной площадке в зависимости от расстояния в северном направлении |
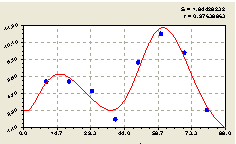
Сравнение показывает, что по аналогии с кварталом № 30, здесь также происходило учащенное воспроизводство взрослой ели в северном направлении от центра. Именно поэтому на лесных плантациях для целлюлозно-бумажных комбинатов ряды посадок ели (аналогично и сосны) ориентируют с юга на север.
Таким образом, северное направление оказалось одинаковым для обеих круговых пробных площадок на разных ельниках. При этом структура модели поведения нисколько не изменилась. Несколько отличаются только значения параметров найденной статистической модели.
Сравнение по коэффициенту корреляции 0.9784 (кв. 37) и 0,9556 (кв. 30) показывает, что увеличение антропогенной нагрузки в действительности расстраивает древостой как систему, то есть популяцию одновидовых или одинаково смешанных видов деревьев. Поэтому коэффициент корреляции является не только статистическим показателем адекватности искомых биотехнических закономерностей по распределениям деревьев на круговой пробной площади, но и становится чувствительным критерием экологического равновесия древостоя.
Северо-восток. В данном геодезическом направлении получили уравнение (рис. 4.31) в виде конструкции
![]()
![]() . (4.29)
. (4.29)
|
Рис. 4.31. График изменения количества деревьев на пробной площадке в зависимости от расстояния в северо-восточном направлении |
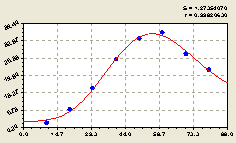
Высокая адекватность полученной биотехнической закономерности по коэффициенту корреляции 0,9962 (0,9931 для кв. № 30) указывает на то, что между лесными деревьями действительно существуют вполне количественно измеримые закономерности в распределении по территории лесного участка.
Из графика на рис. 4.31 видно, что вокруг центральной материнской ели другие деревья расположены неравномерно, но при слабых колебательных возмущениях.
Тогда становится понятным, что расстройство леса (между популяциями деревьев, травы и подлеска, животными, грибами и пр.) и расстройство древостоя (по структуре и видовому разнообразию деревьев) проявляются как сильные и усиливающиеся волновые биотехнические закономерности. Человек первобытный интуитивно ощущал этого, созерцая за поведение лесных и иных деревьев. Ныне цивилизованный человек больше знает только о своих потребностях, чем о потребностях других биологических видов, поэтому и ведет себя как ошалелый от избытка жертвы хищник по басне Крылова о волке в овчарне.
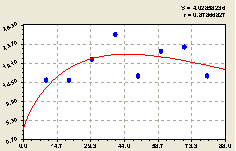
Восток. После проведения структурно-параметрической идентификации получили формулу (рис. 4.32)
![]() . (4.30)
. (4.30)
Высокая адекватность полученной биотехнической модели, конструкция которой оказалась проще по сравнению с формулой (4.3), указывает на существование четкой биотехнической закономерности в распределении всех деревьев на данной территории.
Причем первая составляющая из (4.3) превратилась в формуле (4.30) в постоянный член и тем самым конструкция модели упростилась снижением на два параметра. Однако при этом адекватность повысилась до 0,9955 вместо 0,9190. Отсюда следует, что в разных геодезических направлениях лесной древостой расстраивается по-разному.
|
Рис. 4.32. График изменения количества деревьев на пробной площадке в зависимости от расстояния в восточном направлении |
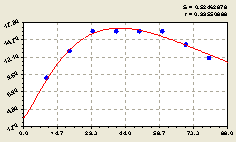
Из графика на рис. 4.32 видно, что в данном геодезическом направлении численность всех деревьев относительно материнской ели изменяется неравномерно по биотехническому закону проф. П.М. Мазуркина.
По формуле (4.30) определили, что количество деревьев, растущих как вблизи, так и далеко от материнской ели, существенно снижается. Поэтому существует оптимальная зона максимального расселения подроста ели от центра обсеменения на расстоянии в пределах 25...60 м.
Юго-восток. По данному направлению получили формулу (рис. 4.33) обратного действия вида
![]() . (4.31)
. (4.31)
|
Рис. 4.33. График изменения количества деревьев на пробной площадке в зависимости от расстояния в юго-восточном направлении |
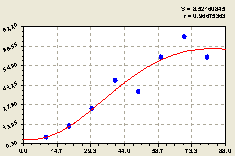
По графику на рис. 4.33 видно, что самое малое количество деревьев наблюдается непосредственно вблизи материнской ели.
То же самое можно подтвердить формулой (4.31), которая показывает снижение численности деревьев.
Подальше от центральной материнской ели происходит колебательное возмущение с растущей амплитудой.
Юг. В ходе проведения структурно-параметрической идентификации по данному направлению получили биотехническую закономерность (рис. 4.34) вида
![]() . (4.32)
. (4.32)
Из графика на рис. 4.35 видно, что численность всех деревьев относительно данного геодезического направления от материнской ели изменяется неравномерно, но волновое изменение вполне четкое.
Об этом свидетельствует волнообразное расположение точек относительно графика.
|
Рис. 4.34. График изменения количества деревьев на пробной площадке в зависимости от расстояния в южном направлении |
Таким образом, в трех геодезических направлениях (восток, юго-восток и юг) по кв. № 37 получили простые, хотя и с двумя составляющими, статистические модели. Простота определяется редукцией первой составляющей из предыдущих формул в постоянный член.
Юго-запад. Как и в северо-восточном направлении, после проведения в этом направлении наблюдается сходство в поведении всех деревьев.
После структурно-параметрической идентификации получили биотехническую закономерность (рис. 4.35) вида
![]()
![]() . (4.33)
. (4.33)
|
Рис. 4.35. График изменения количества деревьев на пробной площадке в зависимости от расстояния в юго-западном направлении |
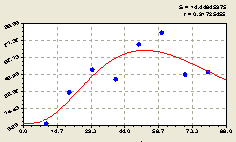
Из графика на рис. 4.35 видно, что по первой составляющей формулы (4.33) происходит снижение численности всех деревьев относительно материнской ели в виде дерева-лидера в зависимости от расстояния L.
В целом, распределение всех деревьев, как и в предыдущих рассмотренных примерах, относительно материнской ели неравномерное и имеет волновой характер.
Запад. В этом направлении получили формулу (рис. 4.36)
![]() . (4.34)
. (4.34)
Из графика на рис. 4.36 видно, что колебательное возмущение численности всех деревьев происходит не близко к центральной ели, как это было на кв. № 30, а подальше от дерева-лидера ели. Поэтому можно вполне предположить, что волновая теория развития и роста лесных деревьев, находящихся в не нарушенных человеком лесорастительных условиях, вполне возможна.
|
Рис. 4.36. График изменения количества деревьев на пробной площадке в зависимости от расстояния в западном направлении |
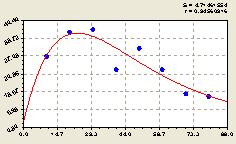
По формуле (4.34) отметим, что на расстоянии 15...30 м от материнской ели находятся больше деревьев, чем ближе или дальше указанного интервала. За пределами 70 м дерево-лидер (трудно было указать, что принятое центральное дерево является материнским деревом) оказывает малое свое влияние на численность всех деревьев, на их рост и развитие, то есть происходит снижение численности всех деревьев относительно центральной ели.
Северо-запад. После проведения в данном геодезическом направлении структурно-параметрической идентификации получили биотехническую закономерность (рис. 4.37) вида
![]()
![]() (4.35)
(4.35)
|
Рис. 4.37. График изменения количества деревьев на пробной площадке в зависимости от расстояния в северо-западном направлении |
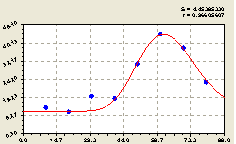
Из графика на рис. 4.37 видно, что на расстоянии 40-80 м от центральной ели произошел рост численности деревьев. Это связано с тем, что центральная ель своей кроной на этом расстоянии уже не затеняет своих соседей и тем самым не влияет на их рост и развитие.
Деревья, которые растут на расстоянии 40-80 м, получают больше тепла и света, чем деревья, растущие непосредственно рядом с материнской елью.
Всего на кольцах. По концентрическим кольцам круговой пробной площади численность всех видов и возрастов деревьев изменяется по формуле биотехнической закономерности
![]()
![]() (4.36)
(4.36)
Графически формула представлена на рис. 4.38.
Из графика на рис. 4.38 видно, что возмущение численности всех деревьев вокруг центральной ели значительно снижается.
|
Рис. 4.38. График изменения общего количества деревьев на кольцевой пробной площадке в зависимости от расстояния |
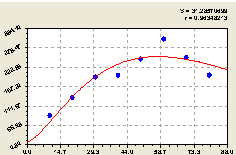
Но колебательное возмущение нарастает по мере удаления деревьев от принятой материнской ели за центр круговой пробной площадки.
Коэффициент корреляции формулы (4.36) стал 0,9635, после усложнения по конструкции по сравнению с моделью (4.27), у которой коэффициент корреляции был гораздо меньше и равен 0,9242.
Таким образом, повышение адекватности модели показывает увеличение экологического равновесия на данный момент времени, то есть текущую устойчивость древостоя по биоэнергетике. При этом усложнение конструкции найденной закономерности может дойти до трехчастной обобщенной биотехнической закономерности.
Тогда можно вполне утверждать не только о компонентном экологическом равновесии древостоя при его достаточной полноте (территориальном равновесии).