
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

4.7. Деревья ели
В табл. 4.5 приведены результаты измерений по численности деревьев ели на круговой пробной площади квартала 37.
Таблица 4.5
Деревья ели, шт.
|
Расстояние от центра |
Геодезические направления |
Всего |
|||||||
|
С |
С-В |
В |
Ю-В |
Ю |
Ю-З |
З |
С-З |
||
|
0 |
3 |
3 |
3 |
3 |
3 |
3 |
3 |
3 |
3 |
|
10 |
6 |
2 |
7 |
3 |
10 |
2 |
19 |
2 |
51 |
|
20 |
6 |
7 |
9 |
4 |
4 |
17 |
23 |
5 |
75 |
|
30 |
5 |
15 |
12 |
16 |
13 |
29 |
23 |
6 |
119 |
|
40 |
2 |
26 |
12 |
19 |
11 |
25 |
14 |
4 |
113 |
|
50 |
8 |
34 |
15 |
27 |
12 |
52 |
23 |
18 |
189 |
|
60 |
11 |
36 |
13 |
50 |
18 |
50 |
18 |
30 |
226 |
|
70 |
9 |
28 |
11 |
52 |
16 |
36 |
11 |
31 |
194 |
|
80 |
3 |
22 |
8 |
38 |
10 |
34 |
9 |
11 |
135 |
|
Итого |
53 |
173 |
90 |
212 |
97 |
248 |
143 |
110 |
1105 |
Север. По данному геодезическому направлению получили уравнение (рис. 4.39) вида
![]()
![]() . (4.37)
. (4.37)
Первая составляющая формулы (4.37) показывает общую тенденцию экспоненциальной гибели (снижения) численности деревьев ели относительно материнского дерева в зависимости от расстояния .
|
Рис. 4.39. График изменения количества деревьев ели на пробной площадке в зависимости от расстояния в северном направлении |
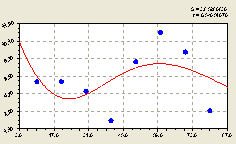
На расстоянии 40 м относительно центрального материнского дерева растет вторая взрослая ель. Очевидно, что по второй составляющей формулы (4.37) резкое волновое возбуждение, что видно из расположения точек около графика на рис. 4.39, численности деревьев ели происходит уже под влиянием второй взрослой ели.
Северо-восток. После проведения структурно-параметрической идентификации уже известной биотехнической закономерности из трех составляющих получили биотехническую закономерность (рис. 4.40)
![]()
![]() (4.38)
(4.38)
|
Рис. 4.40. График изменения количества деревьев ели на пробной площадке в зависимости от расстояния в северо-восточном направлении |
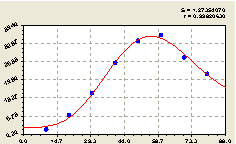
Высокая адекватность биотехнической закономерности указывает на то, что между деревьями ели существует вполне определенная закономерность в распределении на данной территории.
Выявление этой закономерности с волнами, которых здесь не заметно, будет являться целью дальнейших исследований. По графику на рис. 4.40 видно, что недалеко от материнской ели (10...30 м) происходит колебательное возмущение с очень малой амплитудой.
По рис. 4.40 определили, что на расстоянии 60 м численность деревьев ели достигает максимума, а затем идет на спад. Это объясняется тем, что после этого оптимума численность семян от материнской ели резко уменьшается, а вероятность всхожести семян на увеличивающемся расстоянии также снижается. Поэтому на некотором удалении от одной материнской ели должна находиться другая, а затем третья и т.д. Все взрослые семенные ели, которые оказывают негативное влияние на своих соседей, замедляя их рост, развитие и даже вызывая гибель некоторых из них. Но, одновременно, сеть оптимальным образом на земельном участке расположенных семенных деревьев оказывает возбуждение всего древостоя на генерацию новых особей своего вида.
Восток. В данном случае получили формулу (рис. 4.41) вида
![]() . (4.39)
. (4.39)
По графику на рис. 4.41 видно, что близко к центральной ели, как и в северо-восточном направлении, не происходит колебательное возмущение численности деревьев ели. Очень высокий коэффициент корреляции и упрощение конструкции формулы указывают на дружбу между деревьями ели.
|
Рис. 4.41. График изменения количества деревьев ели на пробной площадке в зависимости от расстояния в восточном направлении |
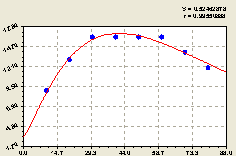
Эта дружба, конечно же, проявляется не через механическое взаимодействие, а через биоэнергетическое общение. Не зря индейцы верили в деревья как в живых существ и не зря марийцы в своей языческой религии обращаются к деревьям в священных рощах.
Юго-восток. После параметрической идентификации предыдущей закономерности получили формулу (рис. 4.42) вида
![]() . (4.40)
. (4.40)
|
Рис. 4.42. Количества деревьев ели на юго-восток |
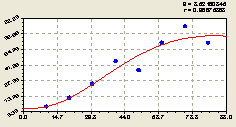
По графику на рис. 4.42 видно, что количество деревьев ели по мере удаления от центрального дерева возрастает. При этом нарастает и волновое возмущение популяции.
Следует отметить, что на данной территории между деревьями ели существуют закономерности распределения, о чем свидетельствует волновое расположение точек на рис. 4.42.
Юг. По данному геодезическому направлению получили биотехническую закономерность (рис. 4.43) вида
![]()
![]() . (4.41)
. (4.41)
Из расположения точек относительно графика на рис. 4.43 видно, что относительно материнской ели среди деревьев ели происходит колебательное возмущение со значительной амплитудой, но на относительно далеком расстоянии от центра.
|
Рис. 4.43. График изменения количества деревьев ели на пробной площадке в зависимости от расстояния в южном направлении |
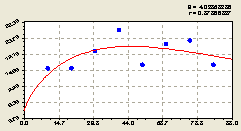
Это связано с волновым расположением точек относительно графика на рис. 4.43. Необходимо отметить, что распределение деревьев ели относительно центрального дерева неравномерное, но с малой амплитудой. На это может оказать влияние направление господствующего ветра, освещенность территории, соседние деревья и др.
Юго-запад. После проведения структурно-параметрической идентификации получили биотехническую закономерность (рис. 4.44) вида
![]()
![]() (4.42)
(4.42)
|
Рис. 4.44. График изменения количества деревьев ели на пробной площадке в зависимости от расстояния в юго-западном направлении |
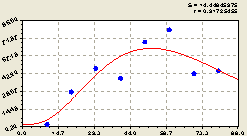
По графику на рис. 4.44 видно, что по мере удаления от центрального материнского дерева количество деревьев ели вначале увеличивается, а затем уменьшается.
При этом на данной территории между деревьями ели наблюдается определенная волновая закономерность в их распределении на лесном участке.
Запад. По биотехнической закономерности (рис. 4.45) вида
![]() (4.43)
(4.43)
получается стрессовое возбуждение численности ели.
|
Рис. 4.45. График изменения количества деревьев ели на пробной площадке в зависимости от расстояния в западном направлении |
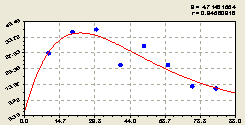
Это, очевидно, связано с тем, что на данной территории растет много взрослых деревьев. Для своего роста и развития они забирают больше света, тепла, питательных веществ. А молодой подрост ели из-за недостатка всех выше перечисленных элементов погибает после 30 м удаления от центрального дерева ели.
Северо-запад. После проведения структурно-параметрической идентификации получили формулу (рис.4.46) вида
![]()
![]() (4.44)
(4.44)
|
Рис. 4.46. График изменения количества деревьев ели на пробной площадке в зависимости от расстояния в северо-западном направлении |
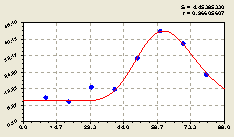
По графику на рис. 4.46 видно, что недалеко от материнского дерева происходит значительное колебательное возмущение численности деревьев ели.
На расстоянии 50-70 м происходит скачок численности деревьев ели.
Всего на кольцах. По концентрическим кольцам круговой пробной площади численность деревьев ели изменяется согласно формуле (4.45)
![]()
![]() . (4.45)
. (4.45)
|
Рис. 4.47. График изменения общего числа деревьев ели на пробной площадке от расстояния |
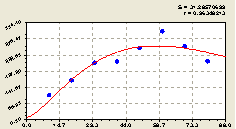
По графику на рис. 4.47 видно, что по мере удаления за 30 м от центрального материнского дерева количество деревьев ели увеличивается, но возрастает и колебательное возмущение популяции.
Высокая адекватность полученной модели (коэффициент корреляции r >0,95) указывает на то, что между деревьями ели существует четкая биотехническая (в смысле мастерства жизни, в данном случае мастерства жизни растущих деревьев ели) закономерность в распределении по территории данного лесного участка.
Кроме того, коэффициент динамичности, как отношение волновых составляющих к основной закономерности в виде не волновых составляющих, должен быть относительно малым.
Повышение коэффициента корреляции и снижение коэффициента динамичности покажут также и о возможной в будущем биологической устойчивости лесных деревьев в данной популяции лесного выдела, то есть данного древостоя в виде ельника.