
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

4.8. Малые деревья ели
Моделирование показывает, что, оказывается, существует биоэнергетика деревьев ели, в виде еще неизвестной волновой субстанции, которая по разным сторонам света различается. Одновидовые деревья, а может и различные симбиотические виды, общаются между собой постоянно, управляя своим развитием и ростом в согласии с другими особями древесных растений.
В табл. 4.6 приведены результаты измерений численности у малых деревьев ели, то есть малого и среднего подроста высотой до 2,0 м, по геодезическим направлениям квартала 30.
Таблица 4.6
Малые деревья ели, шт.
|
Расстояние от центра |
Геодезические направления |
Всего |
|||||||
|
С |
С-В |
В |
Ю-В |
Ю |
Ю-З |
З |
С-З |
||
|
0 |
2 |
2 |
2 |
2 |
2 |
2 |
2 |
2 |
2 |
|
10 |
0 |
0 |
0 |
2 |
7 |
0 |
11 |
0 |
20 |
|
20 |
2 |
0 |
0 |
3 |
4 |
17 |
23 |
2 |
51 |
|
30 |
0 |
0 |
7 |
14 |
12 |
25 |
20 |
0 |
78 |
|
40 |
0 |
0 |
7 |
15 |
8 |
4 |
2 |
0 |
36 |
|
50 |
0 |
7 |
3 |
11 |
5 |
34 |
15 |
0 |
75 |
|
60 |
0 |
14 |
5 |
41 |
15 |
36 |
13 |
0 |
124 |
|
70 |
5 |
10 |
7 |
33 |
10 |
23 |
7 |
16 |
111 |
|
80 |
2 |
10 |
6 |
26 |
7 |
19 |
5 |
8 |
83 |
|
Итого |
11 |
43 |
37 |
147 |
70 |
160 |
98 |
28 |
580 |
Север. После проведения структурно-параметрической идентификации получили биотехническую закономерность (рис. 4.48) вида
![]()
![]() . (4.46)
. (4.46)
По графику на рис. 4.48 видно, что на расстоянии 70 м от центрального дерева происходит резкий рост численности малых деревьев ели.
|
Рис. 4.48. График изменения количества малых деревьев ели на пробной площадке в зависимости от расстояния в северном направлении |
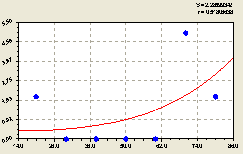
Такой же скачок роста заметен на близком расстоянии от центральной материнской ели.
В среднем по формуле (4.46) характеризуется экспоненциальный рост молодого подроста.
Но молодежи во всех биологических видах очень тяжело пробиться во взрослые особи. Поэтому на расстоянии 20 .. 40 м все проростки ели погибают. Этому мешают взрослые деревья. Поэтому молодняку остается весьма малый шанс на выживание. Они занимают только свободные места в лесном древостое, тем самым повышая территориальное экологическое равновесие при резко волновом изменении по расстоянию от материнской ели компонентное равновесие.
Северо-восток. По данному геодезическому направлению получили формулу (рис. 4.49) вида
![]()
![]() (4.47)
(4.47)
|
Рис. 4.49. График изменения количества малых деревьев ели на пробной площадке в зависимости от расстояния в северо-восточном направлении |
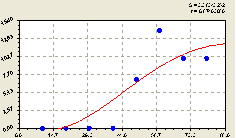
График на рис. 4.49 показывает, что первоначально не наблюдается ни роста, ни гибели молодого подроста. Это связано с тем, что молодые ели на расстоянии 10-40 м не растут.
Можно предположить, что они были не допущены к прорастанию из семян своими соседями. И только на расстоянии 50-60 м происходит резкий скачок роста численности у малых деревьев ели.
Этот факт еще раз показывает высоко напряжение для появления подроста среди взрослых деревьев древостоя.
Восток. После проведения структурно-параметрической идентификации получили биотехническую закономерность (рис. 4.50) вида
![]()
![]() . (4.48)
. (4.48)
|
Рис. 4.50. График изменения количества малых деревьев ели на пробной площадке в зависимости от расстояния в восточном направлении |
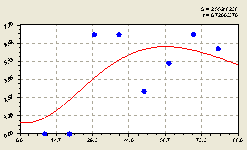
По графику на рис. 4.50 отметим, что распределение молодого подроста ели по данному геодезическому направлению явно неравномерное и притом волновое с затухающим колебанием по мере удаления от центральной материнской ели.
Очевидно, это связано с тем, что на молодой подрост оказывают влияние все взрослые деревья ели и других пород в зависимости от места прорастания семени.
Юго-восток. По данному геодезическому направлению была получена формула (рис. 4.51)
![]()
![]() . (4.49)
. (4.49)
По графику на рис. 4.51 видно, что по мере удаления от центрального материнского дерева происходит постепенный рост численности малых деревьев ели. В данном случае это может быть связано либо с освещением, либо с направлением господствующего ветра и др. при этом заметная четко нарастающая волна возмущения численности подроста по мере удаления от центральной ели.
|
Рис. 4.51. График изменения количества малых деревьев ели на пробной площадке в зависимости от расстояния в юго-восточном направлении |
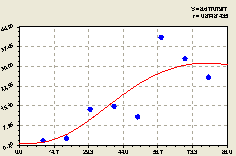
На данном геодезическом направлении наблюдается колебательное возмущение численности молодого подроста ели с растущей амплитудой и убывающим периодом колебания.
Юг. После проведения в данном геодезическом направлении структурно-параметрической идентификации известной конструкции исходной формулы получена биотехническая закономерность (рис. 4.52) вида
![]()
![]() . (4.50)
. (4.50)
Таким образом, во многих направлениях первая составляющая исходной формулы превращается в постоянный член. Стрессовое возмущение в процессах медленной генерации молодняка происходит по биотехническому закону, но, конечно же, только в тех местах лесного древостоя, где нарушается территориальное экологическое равновесие.
|
Рис. 4.52. График изменения количества малых деревьев ели на пробной площадке в зависимости от расстояния в южном направлении |
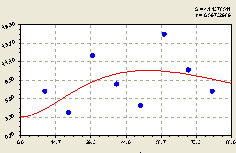
Из рис. 4.52 видно, что молодой подрост ели распределен по лесной территории крайне неравномерно, но только по стрессовому возбуждению самого молодняка. Относительно графика точки четко расположены по волновой закономерности.
Причем амплитуда возмущения нарастает и на это, конечно же, влияет высокая освещенность с южной стороны о материнской ели. Но может оказаться, что на этом месте земельного лесного участка находится зона высокой биоэнергетической активности Земли в виде пучков энергии из глубин мантии.
Это связано с тем, что на рост и развитие молодого поколения оказывают влияние другие взрослые деревья.
Юго-запад. В этом геодезическом направлении получили аналогичную по конструкции формулу (рис. 4.53) вида
![]()
![]() . (4.51)
. (4.51)
|
Рис. 4.53. График изменения количества малых деревьев ели на пробной площадке в зависимости от расстояния в юго-западном направлении |
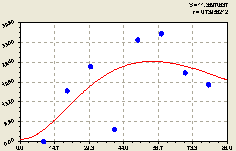
На рис. 4.53 видно, что вблизи материнского дерева происходит сильное колебание численности малых деревьев ели с резко возрастающей к 40 м амплитудой.
Распределение молодого подроста по территории лесного массива, как и в предыдущем примере, крайне неравномерное. При этом на расстоянии 40 м от материнского дерева ели наблюдается явный провал в численности молодняка ели. Здесь растут другие взрослые деревья иных пород. Они оказывают свое непосредственное влияние на торможение роста и развития молодого поколения ели.
Таким образом, в обсеменении лесного участка в данном геодезическом направлении участвуют все взрослые деревья ели. Но их потуги в генерации сдерживаются деревьями иных пород.
Запад. После проведения структурно-параметрической идентификации в этом геодезическом направлении получили биотехническую закономерность (рис. 4.54) вида
![]()
![]() . (4.52)
. (4.52)
|
Рис. 4.54. График изменения количества малых деревьев ели на пробной площадке в зависимости от расстояния в западном направлении |
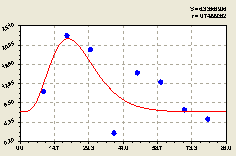
График на рис. 4.54 показывает, что уже вблизи материнского дерева происходит резкий рост численности молодого подроста ели.
На расстоянии 20-80 м наблюдается снижение численности малых деревьев ели. Этот факт также подтвержден формулой (4.52), которая показывает превышение тенденции экспоненциальной гибели молодого подроста в зависимости от расстояния L по сравнению с тенденцией роста популяции.
Северо-запад. В данном направлении получили формулу (рис. 4.55) без первой составляющей в виде уравнения
![]() . (4.53)
. (4.53)
|
Рис. 4.55. График изменения количества малых деревьев ели на пробной площадке в зависимости от расстояния в северо-западном направлении |
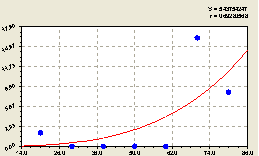
Формула (4.53) показывает тенденцию экспоненциального роста численности молодого подроста относительно центрального материнского дерева в зависимости от расстояния на измеренном участке. Максимум кривой (4.53) наступит гораздо дальше от материнской ели. Причем из графика на рис. 4.55 видно, что резкий скачок роста происходит на расстоянии 60-70 м от центральной ели.
На значительном промежутке от центральной ели численность малых деревцев ели равна нулю. Скачок роста численности малых деревьев ели, очевидно, связан либо с освещенностью данной лесной территории, либо с направлением ветра, который разносит семена от материнской ели и может зависеть от многих других внешних причин.
Всего по кольцам. По концентрическим кольцам круговой пробной площади численность деревьев ели изменяется по формуле (4.54)
![]() . (4.54)
. (4.54)
|
Рис. 4.56. График изменения количества малых деревьев ели на кольцевой пробной площадке |
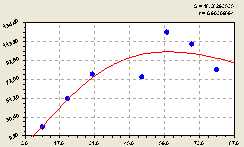
На рис. 4.56 видно, что вблизи материнской ели численность молодого подроста увеличивается уже на расстоянии 10 м.
Это связано либо с освещением территории (молодые ели получают больше тепла и света для своего роста и развития) либо с тем, что при распространении семян ветром наибольшее их количество давно взошло рядом с материнской елью. Поэтому взрослые ели тормозят всхожесть упавших семян.
Таким образом, распределение подроста среди взрослых деревьев происходит с высокой динамичностью расселения. Именно волновая теория здесь правомерно выдвигается на первый план научных исследований, потому что волновые составляющие, в особенности так называемые «импульсные функции», получают наибольшую значимость по сравнению с не волновыми тенденциями.
Поэтому целью дальнейших исследований на множестве круговых пробных площадей становится выявление сложных по конструкции биотехнических закономерностей, включающих уравнения основных тенденций и волновых колебательных возмущений с переменными амплитудой и частотой. Полученные таким образом амплитудно-частотные характеристики (АЧХ) древостоя, по результатам измерений на круговых пробных площадях, позволят количественно косвенно измерять и биоэнергетические поля внутри лесного древостоя.
Кроме того, становится апостериорно, то есть с появлением новых примечательных особенностей поведения деревьев на круговой пробной площади, понятным, что территориальное и компонентное балансы взаимосвязаны. Подрост дает повышение экологического равновесия.