
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

4.12. Малые деревья ели на пробных площадях кварталов 30 и 37
Если древостой изреженный, то появляется территориальное экологическое неравновесие между местами обитания отдельных лесных деревьев. Микро популяции других обитателей леса вообще исчезают. Поэтому для эффективного биоэнергетического взаимодействия нужна оптимальный территориальный баланс в древостое между деревьями разных видов и, конечно же, одного преобладающего вида растений.
Территориальный экологический баланс определяется полнотой древостоя. При снижении этого экологического показателя, изменяющегося в каждой биогруппе деревьев, у деревьев появляется возможность изменения компонентного баланса. Поэтому территориальный и компонентный экологический баланс являются взаимно зависимыми биофизическими величинами.
По-видимому, со снижением полноты древостоя до оптимума, которым является золотая пропорция (будет доказано в конце шестой главы), происходит значимое увеличение численности подроста.
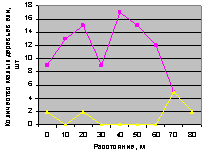
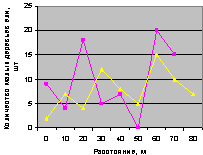
Север и северо-восток. Это наглядно видно на рис. 4.75 и рис. 4.76.
Особенно заметно превышение численности подроста на выделе кв. 30 по сравнению с выделом на кв. 37. Но в конце концентрических колец на расстоянии 70 м оба графика приобретают одинаковое значение.
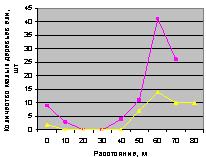
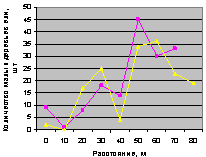
Восток. Десинхроноз в численности подроста по рис. 4.77 на кв. 30.
Юго-восток. Здесь оба графика по рис. 4.78 синхронны. И численность подроста на обоих кварталах леса меняется плавно.
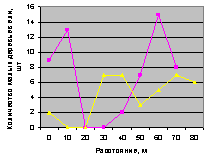
Юг и юго-запад. Графики показаны на рис. 4.79 и рис. 80.
|
Рис. 4.75. Графики изменения количества деревьев ели в зависимости от расстояния (направление север) |
Рис. 4.76. Графики изменения количества деревьев ели в зависимости от расстояния (направление северо-восток) |
|
Рис. 4.77. Графики изменения количества деревьев ели в зависимости от расстояния (направление восток) |
Рис. 4.78. Графики изменения количества деревьев ели в зависимости от расстояния (направление юго-восток) |
|
Рис. 4.79. Графики изменения количества деревьев ели в зависимости от расстояния (направление юг) |
Рис. 4.80. Графики изменения количества деревьев ели в зависимости от расстояния (направление юго-запад) |
Резкое колебание численности подроста повторяет возмущение части популяции в идее взрослых особей. Взаимное распределение всех деревьев и двух поколений ели будет рассмотрено в следующем разделе этой главы. Относительное сопоставление молодняка с взрослым особями по численности может дать новые экологические критерии для оценки качества лесных массивов, находящихся не только на особо охраняемых территориях.
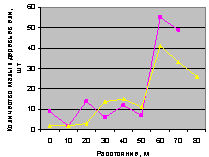
Запад. Здесь происходит сильное колебательное возмущние численности подроста, что видно из рис. 4.81.
|
Рис. 4.81. Графики изменения количества деревьев ели в зависимости от расстояния (направление запад) |
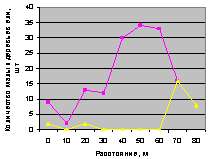
Рис. 4.82. Графики изменения количества деревьев ели в зависимости от расстояния (направление северо-запад) |
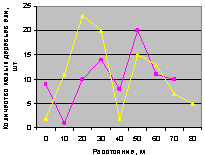
Это действительно связано с волновой природой развития и роста популяции деревьев ели, так как антропогенное влияние рубками деревьев с этой стороны было минимальным.
Северо-запад. Здесь по рис. 4.82 заметны противопоставления двух графиков. На выделе кв. 37 по нижней ломаной линии происходит стрессовый всплеск молодняка по численности только в конце кольцевых пробных площадок. Поэтому этот процесса является естественным и на расстоянии 0...60 м от семенной ели наблюдается ровная популяция молодых деревьев ели.
Совершенно по-иному ведет себя молодняка на выделе кв. 30. Возраст у молодых деревьев ели составляет не более 20 лет.
Иначе говоря, они появились из семян в лесной почве после создания национального парка и запрета ююбой хозяйственной деятельности, в том числе и по рубкам деревьев, на территории лесного массива. Но здесь до создания парка были незаконные рубки, а с 1996 г. проводились санитарные рубки деревьев и рубки для ландшафтного переформирования древостоя (в соседних выделах).
Поэтому биотехнические меры также способствовали снижению территориального баланса и тем самым стремлению ельника к достижению своего компонентного баланса за счет повышенной генерации подроста из семян.
Вполне логично, что даже при одинаковом факеле рассеивания семян от центральной материнской ели больше будет всходов там, где полнота, то есть сомкнутость крон взрослых деревьев, будет меньше.
Таким образом, нужно научиться измерять и точечную полноту.
Подрост ели на кольцевых площадках. На рис. 4.83 показан график распределения подроста по кольцевым пробным площадкам.
|
Рис. 4.83. Графики изменения общего количества малых деревьев ели на кольцевых пробных пощадках от материнской ели |
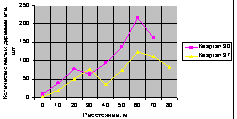
Здесь пики стрессового возбуждения молодого подроста ели по двум кварталам находятся на одном расстоянии 60 м от материнского дерева. Причем на расстоянии 20...60 м на выделе кв. 30 и 30...60 м выдела на кв. 37 наблюдается спад молодого подроста.