
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

4.13. Распределение деревьев на пробных площадях
На рисунках этого раздела обозначения графиков приведены внизу подрисуночной подписи к рис. 4.84 и рис. 4.85.
Мы уже знаем, что любой показатель в любом геодезическом направлении на любом лесном выделе имеет одну единственную общую конструкцию биотехнической закономерности. В частных случаях из трех составляющих остается две или даже одна.
Поэтому параметры модели по тем составляющим и компонентам отдельной составляющей модели, которые исключились в ходе структурной и параметрической идентификации, получают значение нуль или единица. Это позволит в будущем проводить автоматическую идентификацию обобщенной биотехнической закономерности.
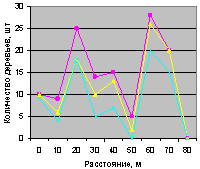
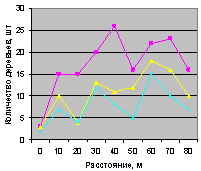
Север на кварталах 30 и 37 показан на рис. 4.84 и рис. 4.85. Сравнение возможно по трем группам деревьев.
Все три группы деревьев (все, ель, малая ель) на северном направлении внутри выдела кв. 30 одинаковы по характеру изменения в зависимости от расстояния от материнской ели. На квартале 37, хотя на расстоянии 40 м наблюдается спад численности всех деревьев и деревьев ели, молодняк ели не появился. Это место произрастания требует к себе повторного внимания, так как из девяти деревьев только два являются взрослой елью. В чем причина не появления всходов ели, хотя вроде бы здесь мала полнота ельника? Может быть, здесь оказывает влияние животный мир ельника? Например, имеются муравейник, или др.
|
Рис. 4.84. Графики изменения количества деревьев ели в зависимости от расстояния (направление север, квартал 30) |
Рис. 4.85. Графики изменения количества деревьев ели в зависимости от расстояния (направление север, квартал 37) |
||
|
|
|
|
|
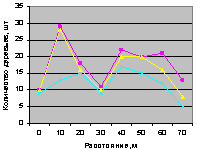
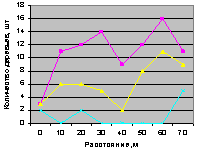
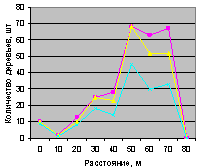
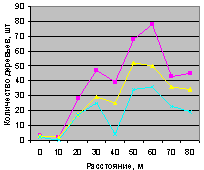
Северо-восток на кварталах 30 и 37. На рис. 4.86 для кв. 30 видно, что все деревья, ель и молодняк ели распределяются одинаково.
|
Рис. 4.86. Количество деревьев ели (северо-восток, квартал 30) |
Рис. 4.87. Количествоа деревьев ели (северо-восток, квартал 37) |
А вот на выделе кв. 37 столь много деревьев, из них преобладает численность взрослой ели, что территориальный баланс уже достигнут, и поэтому уже с расстояния 10 м до 40 м нет места для подроста ели. Но неизвестно, а почему в интервале расстояний 50 .. 70 м, когда и так много стало растущих деревьев, наблюдается резкий скачок подроста ели.
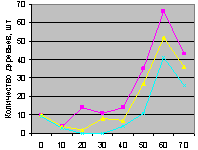
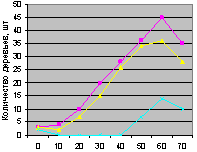
Восток на кварталах 30 и 37. Графики есть на рис. 4.88 и рис. 4.89.
Снова синхронно ведут себя поколения ели на выделе кв. 30 и несколько асинхронно - на выдел кв. 37. первое можно объяснить только искусственным снижением полноты древостоя в соответствующих геодезических направлениях. А второе требует дальнейших исследований.
|
Рис. 4.88. Количество деревьев ели (В, кв. 30) |
Рис. 4.89. Количество деревьев ели (В, кв. 37) |
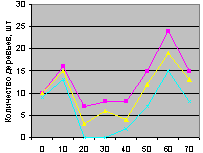
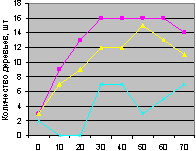
Юго-восток на кварталах 30 и 37 показан на рис. 4.90 и рис. 4.91.
|
Рис. 4.90. Графики изменения количества деревьев ели в зависимости от расстояния (юго-восток, квартал 30) |
Рис. 4.91. Графики изменения количества деревьев ели в зависимости от расстояния (юго-восток, квартал 37) |
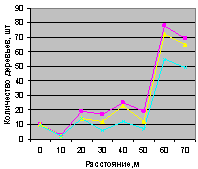
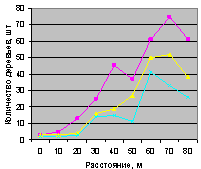
В отличие от синхронности на выделе кв. 30 в мало тронутом древостое выдела кв. 37 снова наблюдается небольшая асинхронность между деревьями ели и всеми деревьями. Но между деревьями ели и подростом ели наблюдается хорошее согласие по численности.
Юг на кварталах 30 и 37. Графики есть на рис. 4.92 и рис. 4.93.
Синхронность поведения деревьев на выделе кв. 30 даже поражает воображение, столь они одинаковы.
Юго-запад на кварталах 30 и 37. Графики на рис. 4.94 и рис. 4.95 показывают хорошую синхронность внутри разновозрастного ельника.
При этом слаженность поведения популяции деревьев одинакова на обоих лесных кварталах.
Запад на кварталах 30 и 37. На рис. 4.96 и рис. 4.97 снова видно слаженное поведение лесных деревьев на обоих кварталах по тем выделам, на которых было малое антропогенное воздействие извне.
|
Рис. 4.92. Графики изменения количества деревьев ели в зависимости от расстояния (направление юг, квартал 30) |
Рис. 4.93. Графики изменения количества деревьев ели в зависимости от расстояния (направление юг, квартал 37) |
|
Рис. 4.94. Графики изменения количества деревьев ели в зависимости от расстояния (юго-запад,квартал 30) |
Рис. 4.95. Графики изменения количества деревьев ели в зависимости от расстояния (юго-запад, квартал 37) |
|
Рис. 4.96. Графики изменения количества деревьев ели в зависимости от расстояния (запад, квартал 30) |
Рис. 4.97. Графики изменения количества деревьев ели в зависимости от расстояния (запад, квартал 37) |
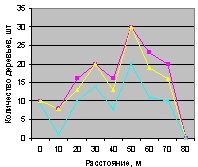
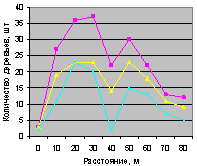
Этот факт еще показывает, что для проведения в дальнейшем других экспериментов не обязательно измерять численность деревьев на всех видах пробных площадок. По-видимому, достаточно будет принять прямоугольные площадки по четырем сторонам света. Так нужно поступить с крестообразными пробными площадями на всех выделах лесного кв. 30.
Северо-запад на квартале 30 показан на рис. 4.98 и рис. 4.99.
|
Рис. 4.98. Графики изменения количества деревьев ели в зависимости от расстояния (северо-запад, квартал 30) |
Рис. 4.99. Графики изменения количества деревьев ели в зависимости от расстояния (северо-запад, квартал 37) |
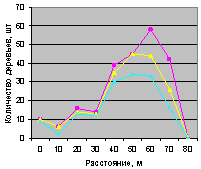
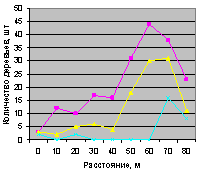
Во всех примерах соблюдалась иерархия численности:
а) численность ели была не больше численности всех деревьев;
б) численность молодняка ели не превышает числа взрослых.
Тогда, для упрощения проведения полевых экспериментов, можно принять только один из показателей, например, общая численность всех деревьев на прямоугольной пробной площадке на крестообразной пробной площади, ориентированной по сторонам света или же в сторону преобладающих ветров сезон рассеивания семян. В итоге получается многообразие технических решений по патентоспособным способам.
Всего на кольцах квартала 30. Синхронизация поведения каждой части популяции всех лесных деревьев видна из рис. 4.100 и рис. 101.
В дальнейшем на земельном участке нужно посмотреть и оценить качество растущих деревьев по сортности ствола, качеству надземной кроны и распластанных по лесной почве корней. С понижением качества растущих особей подрост ели вполне может оживиться по численности. Это означает, что на компонентное равновесие влияет не только недостаток до оптимума территориального баланса в древостое, но жизнеспособность деревьев и их групп.
|
Рис. 4.100. Графики изменения общего количества деревьев ели в зависимости от расстояния (квартал 30) |
Рис. 4.101. Графики изменения общего количества деревьев ели в зависимости от расстояния (квартал 37) |
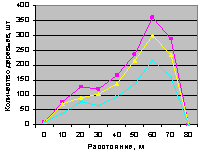
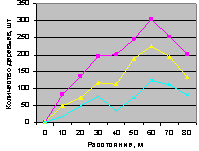
Конечно же, синхронность поведения популяции лесных деревьев на выделе кв. 30 - это результат правильного ухода за лесом со стороны лесных работников национального парка за последние 15...20 лет.
Но ничуть не хуже ведется синхронизация численности популяции у ели на выделе кв. 37. Поэтому, по большому счету, лес не нуждается в уходе со стороны человека. Лишь бы он не мешал лесу, прежде всего его ядру в виде древостоя, самостоятельно развиваться и расти. Даже после достижения биологической старости лесные деревья не теряют, а наращивают, свою экологическую ценность. В связи с этим климаксовые леса гораздо ценнее обилием внутренних экосистем по сравнению с технически спелой древесиной.
Но одно ясно, - нужно понять закономерности поведения каждого лесного выдела и помочь ему уменьшить свои волновые возмущения снижением с помощью «лесного человека» амплитуды колебания.