
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

5.4. Доля малых деревцев ели среди деревьев ели
В табл. 5.3 приведены результаты измерений доли (можно было указать и в процентах) у малых деревцев ели среди деревьев ели на выделе 9 лесного квартала № 30.
Таблица 5.3
Доля малых деревцев ели среди деревьев ели ![]()
|
Расстояние от центра |
Геодезические направления |
Всего |
|||||||
|
С |
С-В |
В |
Ю-В |
Ю |
Ю-З |
З |
С-З |
||
|
0 |
0.9000 |
0.9000 |
0.9000 |
0.9000 |
0.9000 |
0.9000 |
0.9000 |
0.9000 |
0.9000 |
|
10 |
0.4643 |
0.7500 |
0.8667 |
1.0000 |
0.6667 |
0.5000 |
0.1250 |
0.3333 |
0.5493 |
|
20 |
0.9375 |
0.0000 |
0.0000 |
1.0000 |
1.0000 |
0.8000 |
0.7692 |
0.9286 |
0.8667 |
|
30 |
0.9000 |
0.0000 |
0.0000 |
0.5000 |
0.5000 |
0.7200 |
0.7000 |
0.9231 |
0.6154 |
|
40 |
0.8500 |
0.5714 |
0.5000 |
0.5217 |
0.5385 |
0.6087 |
0.6154 |
0.8571 |
0.6812 |
|
50 |
0.7500 |
0.4074 |
0.5833 |
0.5833 |
0.0000 |
0.6618 |
0.6667 |
0.7556 |
0.6435 |
|
60 |
0.7500 |
0.7885 |
0.7895 |
0.7639 |
0.7692 |
0.5769 |
0.5789 |
0.7500 |
0.7233 |
|
70 |
0.6250 |
0.7222 |
0.6154 |
0.7538 |
0.7500 |
0.6346 |
0.6250 |
0.6154 |
0.6864 |
|
Итого |
0.7422 |
0.6438 |
0.6585 |
0.7333 |
0.7429 |
0.6529 |
0.6434 |
0.7720 |
0.6884 |
На квартале № 30 как вблизи материнской ели, так и на некотором расстоянии от нее, произрастает большое количество деревьев ели. Среди них выделяется много малых деревцев ели.
Таким образом, доля молодого подроста будет изменяться в зависимости от его количества на определенном расстоянии от материнской ели.
В табл. 5.3 доля малых деревьев ели среди деревьев ели принятого древостоя изменяется также от нуля до 1,0000. По-видимому, из-за антропогенного влияния на весь квартал леса № 30, доля подроста среди ели стала неуправляемой со стороны популяции хвойных деревьев. Поэтому устойчивость структуры древостоя снижается.
Север. В этом направлении после структурно-параметрической идентификации получили (рис. 5.19) уравнение
![]()
![]() . (5.19)
. (5.19)
|
Рис. 5.19. График изменения доли малых деревцев ели среди деревьев ели на пробной площадке в зависимости от расстояния в северном направлении |
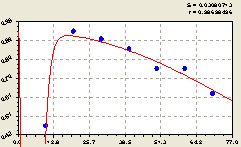
Первая составляющая формулы (5.19) показывает тенденцию экспоненциальной гибели (спада).
Интенсивность гибели (спада) составляет 2,22137, то есть в этом направлении существует угроза полного уничтожения молодого подроста.
Поэтому нужно предусмотреть защитные мероприятия, которые помогут не только остановить гибель молодого подроста, но и будут способствовать их росту и развитию.
К таким мероприятиям относятся рубки деревьев для ухода за лесом, санитарные мероприятия, формирование ландшафта и др.
Северо-восток. После идентификации биотехнического закона получили (рис. 5.20) зависимость
![]()
![]() . (5.20)
. (5.20)
Как и в северном направлении, по первой составляющей формулы (5.20) видно, что снова наблюдается тенденция гибели молодого подроста, а значит, доля малых деревцев ели на этом направлении будет снижаться по мере удаления семян от материнского дерева, располагаемого в экспериментах в центре круговой пробной площади.
|
Рис. 5.20. График изменения доли малых деревцев ели среди деревьев ели на пробной площадке в зависимости от расстояния на северо-восток |
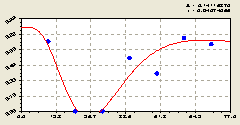
Интенсивность гибели составляет 2,08528, то есть в этом направлении также существует угроза полного уничтожения молодого подроста ели не удалении в интервале 20...30 м.
Здесь также необходимо проводить те же биотехнические мероприятия, которые были указаны в предыдущем разделе книги.
Восток. В этом геодезическом направлении получили (рис. 5.21) уравнение биотехнической закономерности
![]()
![]() . (5.21)
. (5.21)
|
Рис. 5.21. График изменения доли малых деревцев ели среди деревьев ели на восток |
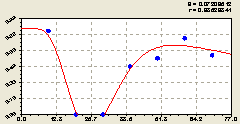
По первой составляющей формулы (5.21) происходит снижение доли молодого подроста ели среди деревьев ели.
Это происходит из-за того, что малым деревьям ели не хватает благоприятных условий для развития и дальнейшего роста.
Они начинают погибать, располагаясь недалеко от материнской ели.
Юго-восток. Здесь получили биотехническую закономерность (рис. 5.22) в виде двухчленного уравнения с волновой составляющей
![]()
![]()
![]() . (5.22)
. (5.22)
Амплитуда колебания имеет максимум, изменяясь по биотехническому закону вдоль юго-востока от центральной ели, а частота колебания возрастает из-за увеличения половины периода колебания.
|
Рис. 5.22. График изменения доли малых деревцев ели среди деревьев ели на пробной площадке в зависимости от расстояния на юго-восток |
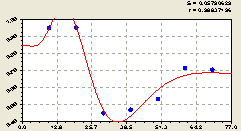
Хотя и принимали решение не углубляться в волновую теорию и соответствующие закономерности колебательного возмущения популяции лесных деревьев, однако в данном направлении коэффициент динамичности размещения подроста ели столь высок, что тренд не дает объективной картины влияния расстояния от материнской ели.
Юг. Для этого направления была получена (рис. 5.23) формула
![]()
![]()
![]() . (5.23)
. (5.23)
|
Рис. 5.23. График изменения доли малых деревцев ели среди деревьев ели на пробной площадке в зависимости от расстояния в южном направлении |
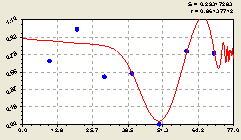
График показывает сдвиг волны с резко учащающейся частотой подальше от материнской ели.
На участке 0...40 м возможна еще одна колебательная волна.
Таким образом, южное направление весьма неоднозначное и сложное для жизнедеятельности появляющихся всходов ели. Конкуренция за свет заставляет подрост и взрослых деревьев ели бороться друг с другом за освещение.
Юго-запад. В этом геодезическом направлении после структурно-параметрической идентификации получили биотехническую закономерность (рис. 5.24) вида
![]()
![]() . (5.24)
. (5.24)
Здесь возмущение заметно лишь вблизи материнского дерева ели, то есть на расстоянии 0...10 м.
|
Рис. 5.24. График изменения доли малых деревцев ели среди деревьев ели на пробной площадке в зависимости от расстояния на юго-запад |
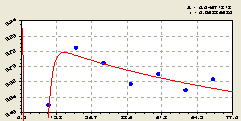
По графику на рис. 5.24 получается, что, в целом, по юго-западному направлению сохраняется общая тенденция снижения доли молодого подроста среди деревьев ели в зависимости от расстояния . При этом популяция в этом секторе круговой пробной площади успокаивается и расположение точек около графика показывает малую динамичность волнового возмущения ели.
Запад. По этому направлению в результате идентификации биотехнического закона было получено (рис. 5.25) уравнение
![]()
![]() . (5.25)
. (5.25)
|
Рис. 5.25. График изменения доли малых деревцев ели среди деревьев ели на пробной площадке в зависимости от расстояния в западном направлении |
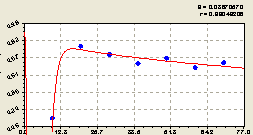
Первая составляющая формулы (5.25) указывает на снижение доли малых деревцев ели среди деревьев ели по закону экспоненциальной гибели (спада) с интенсивностью гибели, равной 0,60945.
На расстоянии 20...70 м происходит незначительное снижение доли молодого подроста по второй составляющей. Но кризис наступает только на 0...20 м.
Северо-запад. С увеличением расстояния от центрального дерева-лидера, то есть материнской ели, доля малых деревцев ели среди всех учтенных деревьев ели изменяется (рис. 5.26) по формуле
![]()
![]() . (5.26)
. (5.26)
|
Рис. 5.26. График изменения доли малых деревцев ели среди деревьев ели на пробной площадке в зависимости от расстояния в северо-западном направлении |
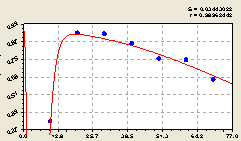
Здесь также произошел кризис молодого поколения. На это указывает отрицательный знак перед второй составляющей статистической модели (5.26). На кризис, по-видимому, больше влияет освещенность, но также и сильная биоэнергетика матери, так как здесь света мало.
Как и в западном направлении, здесь также наблюдается общая тенденция снижения доли молодого подроста среди всех деревьев ели.
Поэтому необходимо проведение мероприятий, которые позволят молодому подросту активно развиваться.
Это, например, вырубка старых засохших и больных лесных деревьев, мероприятия по очистке лесной территории от ненужного хлама, мусора и т.д.
Все такие меры относятся к природоприближенному обустройству территорий, в нашем случае национального парка.
Вместе с тем, исходя из принципа «лес не нуждается в уходе», можно выявить, по крайней мере, следующие силы действия на подрост ели, то есть на свое ежегодное потомство:
а) прямое положительное действие, по всем кольцевым пробным площадкам, центральной материнской ели на молодняк своего вида из своих семян, которое медленно убывает с расстоянием и. по-видимому, в репродуктивном возрасте простирается это биоэнергетическое действие более чем на 100 м;
б) в секторе от юго-запада до севера, то есть с углом в 1350, на близком расстоянии от себя материнская ель сама подавляет появление елового подроста, доводя долю молодых деревьев ели около себя до нуля;
в) в угловом секторе 900 от севера до востока процесс подавления матерью своих отпрысков распространяется дальше от себя, и материнская ель доводит почти до нуля долю своего подроста на части кольца радиусом 20...60 м;
г) на территории круговой пробной площади в секторе от востока до юго-запада, с углом в 1350, биоэнергетическое действие материнской ели приходит в сильное колебательное возмущение от влияния солнечного освещения, то есть солнечной энергии.
Известно, что дерево качаясь на ветру, работает как водяной насос, поднимая по капиллярным пустотам клеток с устьицами в виде обратных клапанов, питательную жидкость от корней к листве (хвое). поэтому на радиальное распределение подроста влияет, по крайней мере, силы от трех источников: биоэнергетика от материнской ели, световая энергия Солнца и механическая энергия от преобладающих ветров. влияет ли энергия лунного прилива и отлива, как это заметно на животных, мы пока не знаем. Но то, что клетки паренхимы и ксилемы между корой и заболонью древесины имеют цикл возникновения, роста и созревания происходит в течение семи суток - известно из биологии растений, например из книги [].
Всего на кольцах. По концентрическим кольцам круговой пробной площади доля малых деревцев ели среди всех деревьев ели изменяется (рис. 5.27) по простой формуле экспоненциальной гибели
![]() . (5.27)
. (5.27)
|
Рис. 5.27. График изменения доли малых деревцев ели среди деревьев ели на пробной площадке в зависимости от расстояния |
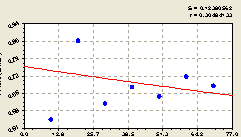
Формула (5.27) показывает снижение доли молодого подроста ели среди деревьев ели, причем четко по закону экспоненциальной гибели, по мере удаления подроста от материнской ели.
Таким образом, доля малых деревцев ели среди деревьев ели по всем направлениям снижается. Как видно из расположения точек около графика, это плавное снижение сопровождается колебательным возмущением, ослабляющимся по амплитуде по мере удаления от материнской ели. Заметно также, что частота колебания снижается и это биоэнергетическое колебательное возмущение успокаивается.
Понятно, что около матери энергия намного превышает потребный для подроста уровень. По азимуту к этому добавляются еще воздействия солнечных лучей, притяжения Луны и раскачивания от ветра. При благоприятных сочетаниях тепла, влажности и питания минеральными веществами материнская ель один раз в 5...7 лет от опыления пыльцой от нижних ветвей с мужскими органами дает в верхней части своей кроны обильные урожаи шишек с семенами.
В итоге даже одно дерево ели способно генерировать потомство, образуя вокруг себя не меньше чем на 100 м отдельные биогруппы из собственных потомков по различным радиальным направлениям.