
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

6.2. Плотность всех деревьев на пробной площади кв. 30
Если бы все пробные площадки были одного размера, то плотность размещения и абсолютная численность лесных деревьев были бы равносильными по биотехническим закономерностям (табл. 6.2).
|
Таблица 6.2 Плотность всех видов деревьев n, шт./ар
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Последняя строка в табл. 6.2 и других последующих табличных моделях показывает среднее (не среднеарифметическое, а средневзвешенное) значение показателя по всем пробным площадкам вместе.
Север. Здесь (рис. 6.2) и во всех других формулах появился постоянный член, показывающий постоянство условий произрастания по биотехнической закономерности вида
![]()
![]() . (6.5)
. (6.5)
|
Рис. 6.2. График изменения плотности всех деревьев на пробной площадке в зависимости от расстояния в северном направлении |
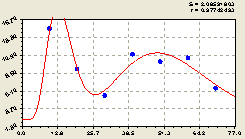
Как видно из расположения точек по фактическим значениям показателя относительно графика на рис. 6.2, вместо двух стрессовых возбуждений можно идентифицировать одно возбуждение на расстояние от материнской ели до 80 м, а затем идентифицировать волновую составляющую колебательного возмущения. Такие биотехнические закономерности покажем для деревьев круговой пробной площадки на выделе лесного квартала 37.
Восток. Получена формула (рис. 6.3) из трех членов в виде
![]()
![]() . (6.6)
. (6.6)
Для хозяйственно измененных лесных участков волновые закономерности трудно получаются из-за анархического поведения людей.
|
Рис. 6.3. График изменения плотности всех деревьев на пробной площадке в зависимости от расстояния в восточном направлении |
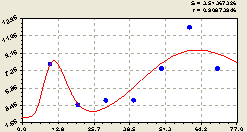
Даже отраслевые специалисты, например лесного хозяйства, до сих пор мало знают о лесной экологии и поведении лесных деревьев и их групп. Их учили считать кубатуру стволов и выращивать лесные насаждения.
Поэтому гораздо труднее доказывать хозяйственникам, в особенности бизнес-элите от кругляка, о закономерностях леса и древостоев.
|
Рис. 6.4. График изменения плотности всех деревьев на пробной площадке в зависимости от расстояния в южном направлении |
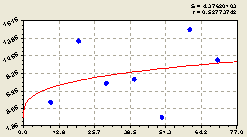
Юг. Как видно из графика на рис. 6.4, здесь тренд получился упрощением (редукцией) биотехнического закона до показательного закона ![]()
При этом с постоянным членом, снова показывающим свойство места произрастания около центральной материнской ели и в круге до 100 м от неё, оба стресса превращаются в две волны, первая из которых является недоразвитой.
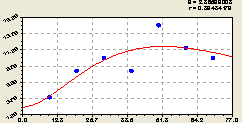
Запад. По графику на рис. 6.5 видно, что по формуле
![]() (6.8)
(6.8)
По сравнению с другими сторонами света наблюдается спокойная динамика плотности всех деревьев на пробных площадках прямоугольной формы.
Причины такого поведения популяции лесных деревьев пока неизвестны.
Однако понятно, что коэффициент динамичности на западном направлении будет наименьшим.
Всего на кольцах. По рис. 6.6 в соответствии с закономерностью
![]()
![]() (6.9)
(6.9)
|
Рис. 6.5. График изменения плотности всех деревьев на пробной площадке в зависимости от расстояния в западном направлении |
|
Рис. 6.6. График изменения плотности всех деревьев на кольцевой пробной площадке от расстояния |
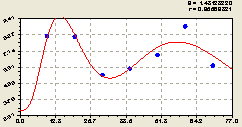
наблюдается картина изменения плотности всех видов деревьев на круговой пробной площади по аналогии с направлениями север и восток. Тем самым западное направление как бы компенсирует сильную флюктуацию плотности лесных деревьев на южном направлении. Таким образом, становится понятным, что волновая динамика происходит не только по радиусам круговой пробной площади, но и по кольцам.
Интересным является тот факт, что, по расположению точек около графика на рис. 6.6, волна небольшой длины наблюдается только на участке расстояния от материнской ели в пределах 50...70 м.
Этот факт показывает, что колебание по кольцам происходят как круги на водной поверхности, однако эти круги начинаются не от места падения камешка на воду, а гораздо дальше.
Пространственная картина (рис. 6.7) наглядно показывает совместное влияние радиального расстояния и азимута центров пробных прямоугольных и секторных пробных площадок на плотность размещения лесных деревьев.
На двухфакторном графике на рис. 6.7, построенном в программной среде TableCurve 3D по измеренным и вычисленным данным плотности территориального распределения деревьев, заметны несколько возвышений.
Эти «горы» и есть места нахождения биогрупп лесных деревьев.
Картина распределения показывает, что около материнской ели находятся две биогруппы, а на расстоянии 50...70 м по всей окружно сти находятся уже четыре биогруппы лесных деревьев.
|
Рис. 6.7. Изменение плотности всех деревьев на пробной площади от расстояния и азимута |
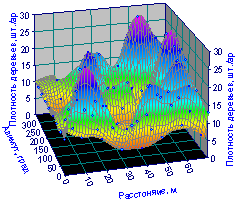
Но частота появления биогрупп деревьев убывает с расстоянием от центра из-за того, что на радиусе в пределах 20...30 мм, при длине окружности 2ΠL = 2Π x 25 = 50 Π, плотность размещения биогрупп будет равна ![]() . А на окружности со средним радиусом 60 м этот параметр симбиотической структуризации древостоя будет равным
. А на окружности со средним радиусом 60 м этот параметр симбиотической структуризации древостоя будет равным
![]() .
.
Отношение плотностей распределения биогрупп лесных деревьев будет равным, при переходе от радиуса 25 м к радиусу 60 м от центральной материнской ели, ![]() . Тогда можем составить пропорции вида: 1 / 0, когда в центре круговой пробной площади находится сама материнская ель; 2 /25 на среднем расстоянии 25 м расположены две биогруппы; 4 / 60 на среднем расстоянии 60 м от материнской ели расположены четыре биогруппы лесных деревьев.
. Тогда можем составить пропорции вида: 1 / 0, когда в центре круговой пробной площади находится сама материнская ель; 2 /25 на среднем расстоянии 25 м расположены две биогруппы; 4 / 60 на среднем расстоянии 60 м от материнской ели расположены четыре биогруппы лесных деревьев.
По эти трем точкам получится биотехническая закономерность изменения количества NБГД групп лесных деревьев в виде ориентировочного математического выражения ![]() с коэффициентом корреляции, равным единице.
с коэффициентом корреляции, равным единице.