
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

6.5. Плотность всех деревьев на пробной площади кв. 37
Табличная модель приведена в данных табл. 6.5.
|
Таблица 6.5 Плотность всех видов деревьев n , шт./ар
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Результаты предыдущего анализа показали, что круговая пробная площадь на выделе 8 кв. 37 является более экологически сформированной и гармоничной по структуре по сравнению с выделом 9 кв. 30.
Причем заметим, что доля ели на выделе 8 кв. 37 равна 50 % по сравнению с выделом 9 на кв. 30. Поэтому и территориальная структуризация, т. е. территориальное экологическое равновесие (баланс), у деревьев ели здесь богаче. Поэтому будем искать сразу же волновые закономерности.
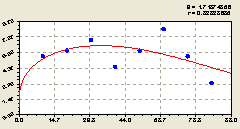
Север. Основная тенденции, которая называется трендом, показана на рис. 6.20 после идентификации не волновых составляющих формулы
![]() . (6.20).
. (6.20).
|
Рис. 6.20. Тренд изменения плотности всех деревьев на пробной площадке в зависимости от расстояния в северном направлении |
Здесь появились две составляющие.
Самым основным является постоянный член, который появился после упрощения первой составляющей в виде закона экспоненциальной гибели, часто приведенной в предыдущих главах книги.
Он показывает, как уже указывалось в предыдущих примерах данной главы, биоэнергетическую потенцию места произрастания всей круговой пробной площади. Соответственно, как и все лесоводы, будем расширенно понимать этот потенциал в среднем для всего лесного выдела.
По остаткам от полученной формулы (6.20) после моделирования параметрической идентификацией волнового уравнения асимметричного вейвлет-сигнала (рис. 6.21) было получено волновое уравнение вида
![]()
![]() . (6.21)
. (6.21)
Перед cos находится половина амплитуды, в внутри - полупериод, поэтому функция (6.21) имеет амплитудно-частотную характеристику.
|
Рис. 6.21. График волны изменения плотности всех деревьев на пробной площадке по остаткам в зависимости от расстояния в северном направлении |
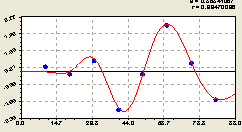
Отрицательный знак перед формулой (6.21) показывает, что этот вейвлет-сигнал указывает на кризис в поведении популяции всех деревьев. Иначе говоря, в северном направлении лента деревьев шириной 20 м испытывает дискомфорт в биоэнергетических отношениях. При этом коэффициент корреляции столь высокий, что такие выводы о поведении сообщества лесных деревьев становятся однозначными и вполне уместными.
Вместе функциональную математическую модель (рис. 6.22) получили, после совместной параметрической идентификации, в виде
![]()
![]()
![]() . (6.22)
. (6.22)
|
Рис. 6.22. График изменения плотности всех деревьев на пробной площадке по остаткам в зависимости от расстояния на север |
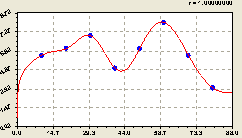
Коэффициент корреляции равен единице.
Процесс совместной параметрической идентификации двух раздельно полученных составляющих формулы (6.22) называется «встряской», что по аналогии похоже на уплотнение различных предметов в ящике путем встряхивания.
Восток. На рис. 6.23 показан график тренда по уже знакомой формуле
![]() . (6.23)
. (6.23)
Здесь по точкам вокруг основной тенденции сразу же заметна малая динамичность процесса биоэнергетического взаимодействия между лесными деревьями. Но параметры динамичности, из-за малого места данной книги, здесь не рассматриваются.
Можно только отметить, что отношение второй составляющей формулы типа (6.22) к не волновым составляющим готовой модели показывает приспособляемость древостоя к среде.
|
Рис. 6.23. Тренд изменения плотности всех деревьев на пробной площадке в зависимости от расстояния в восточном направлении |
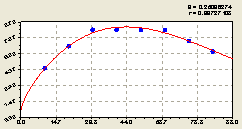
Практически можно считать, что на восточном направлении на ленте шириной в 20 м все деревья расположились гармонично и без волновых возмущений биоэнергетики меду собой.
Однако даже такое малое колебательное возмущение можно измерить вейвлет-функцией (рис. 6.24) с переменой амплитудой и частотой вида
![]()
![]() . (6.24)
. (6.24)
|
Рис. 6.24. График волны изменения плотности всех деревьев на пробной площадке по остаткам в зависимости от расстояния на восток |
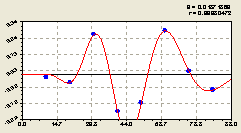
Программная среда CurveExpert - 1.3 каждый раз показывает график с расширением даже малых ординат на всю высоту окна показа графика.
И это становится удобным и наглядным для последующего анализа на основе апостериорной информации, то есть информации, появляющейся в процессе осмысления хода моделирования.
Вместе, как показано на рис. 6.25, после объединения двух составляющих получилась аналогичная северному направлению формула
![]()
![]()
![]() . (6.25)
. (6.25)
Эвристической идентификацией найденных свойств и отдельных закономерностей, то есть апостериорной информации, формулируются новые выводы и выдвигаются неизвестные гипотезы.
|
Рис. 6.25. График изменения плотности всех деревьев на пробной площадке в зависимости от расстояния на восток |
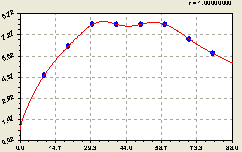
В этой книге эвристические приемы не приводим, а также не показываем также и результаты эвристического анализа параметров модели, усложняющейся в виде последующих составляющих до конструкции готовой статистической модели.
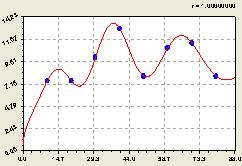
Юг. График на рис. 6.26 и биотехническая закономерность показывают плавное изменение плотности лесных деревьев по формуле
![]() . (6.26)
. (6.26)
|
Рис. 6.26. Тренд изменения плотности всех деревьев на пробной площадке в зависимости от расстояния в южном направлении |
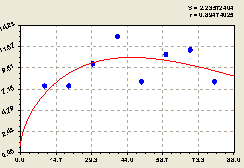
Она показывает тренд по влиянию расстояния.
Можно узнать и время процесса отношений между деревьями, если измерять у каждого из них возраст. Также можно определять и скорость протекания процессов симбиотического развития и роста по отдельным особям лесного древостоя, на территории круговой пробной площади.
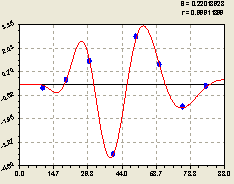
По остаткам (рис. 6.27) получилась вейвлет-фцункция вида
![]()
![]() . (6.27)
. (6.27)
В знаменателе под функцией косинуса находится формула половины периода колебания. Она показывает, что около материнского дерева полная длина волны была равной 2 x 11,6927 ≈ 23,4 м.
В итоге получается, что длина биоэнергетической волны материнской ели равна примерно её высоте. По мере удаления от центрального дерева ели длина волны нарастает, поэтому частота колебательного возмущения снижается.
|
Рис. 6.27. График волны изменения плотности всех деревьев на пробной площадке по остаткам в зависимости от расстояния на юг |
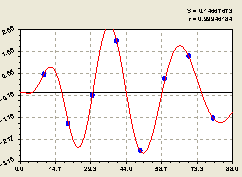
Этот факт указывает на то, что популяция деревьев в южном направлении успокаивается по мере удаления от центра. Кроме успокоения по частоте возмущения, из графика на рис. 6.27 видно, что площади над и под осью абсцисс примерно одинаковы. Это - свойство вейвлета.
Вместе (рис. 6.28) из всех составляющих после параметрической идентификации получается биотехническая закономерность вида
![]()
![]()
![]() . (6.28)
. (6.28)
|
Рис. 6.28. График изменения плотности всех деревьев на пробной площадке от расстояния в южном направлении |
Как было указано в предыдущих главах книги, важна не наличие сильного биоэнергетического колебания, а его результативный характер, то есть амплитудно-частотная характеристика волнового возмущения.
Но, как и по предыдущим сторонам света, на юг колебательное возмущение имеет кризисный для популяции лесных деревьев характер.
Запад. Здесь (рис. 6.29), по формуле
![]() , (6.29)
, (6.29)
проявляется успокаивающийся характер поведения популяции деревьев.
|
Рис. 6.29. Тренд изменения плотности всех деревьев на пробной площадке в зависимости от расстояния в западном направлении |
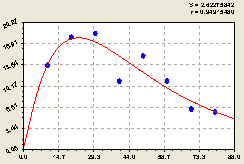
Заметно по расположению точек фактических значений показателя около графика на рис. 6.29, что относительно небольшие волновые циклично-колебательные изменения происходят на расстоянии 20...80 м от центра.
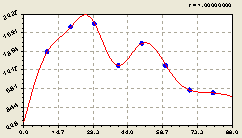
По остаткам (рис. 6.30) получилась формула известной конструкции
![]()
![]() . (6.30)
. (6.30)
|
Рис. 6.31. Волна плотности деревьев по остаткам |
Практически, в условиях проведения лесной инспекции, можно рассчитать коэффициент динамичности даже без выявления последующих колебательных возмущений: просто остатки от формулы типа (6.29) нужно поделить на саму формулу (6.29). А её в виде основой тенденции нетрудно получить даже студентам.
Вместе (рис. 6.32) тренд и волна дают биотехническую закономерность в виде двучленной формулы
![]()
![]()
![]() . (6.31)
. (6.31)
|
Рис. 6.32. График изменения плотности всех деревьев на пробной площадке от расстояния в западном направлении |
Теперь рассмотрим знак перед сдвигом начала волны, то есть в формуле (6.31) число 3,93144. Отрицательный знак показывает, что волновое возмущение получает начало раньше начала системы координат.
Поэтому на западном направлении начало координат биоэнергетического воздействия находится за материнским деревом на восток, причем на расстоянии сдвига 3,93144 9,93375 / 12,43 м.
Отсюда следует, что собственное биоэнергетическое поле материнской ели является асимметричным и достаточно крупным по площади.
Всего на концентрических кольцах (рис. 6.33) лесные деревья распределяются по двучленной формуле
![]() . (6.32)
. (6.32)
Постоянный член можно было поставить как первую составляющую модели. Эту иерархию следует соблюдать при значительном количестве членов модели. На простом уравнении возможна произвольная запись.
|
Рис. 6.33. Тренд изменения плотности всех деревьев на кольцевой пробной площадке от расстояния |
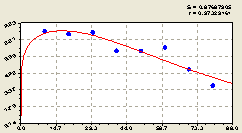
Тренд аналогичен формулам радиальных направлений. Этот факт позволяет применять в измерениях любое геодезическое направление, например, в сторону только преобладающих ветров, или же в сторону лесной прогалины или опушки и т.д.
По остаткам (рис. 6.34) получено уравнение вида
![]()
![]() , (6.33)
, (6.33)
которое одинаково по конструкции с предыдущими моделями.
|
Рис. 6.34. График волны изменения плотности всех деревьев по остаткам на пробной площадке в зависимости от расстояния |
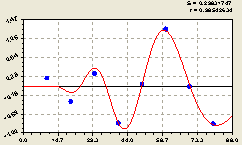
Тогда, при подтверждении предыдущих формул по конструкции, на большом количестве практических примеров из разных мест и условий произрастания, можно назвать обобщенной формулой или же законом распределения лесных деревьев относительно семенных особей.
Вместе (рис. 6.35) все три составляющие дали формулу
![]()
![]()
![]() . (6.34)
. (6.34)
В экспериментах мы полагали, что количество деревьев на малой пробной площадке даст резкие колебания в плотности. Поэтому для крестообразной пробной площади принимали пробные площадки в виде прямоугольников 2BxB.
|
Рис. 6.35. График изменения плотности всех деревьев на пробной площадке в зависимости от расстояния |
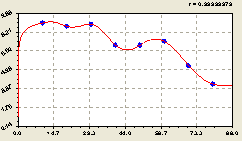
Однако это структурное изменение приводит к нарушению симметрии в центре с получением размеров квадрата 20 x 20 м.
В итоге удельный территориальный показатель плотности размещения деревьев становится мало сопоставимым с первичным показателем по численности лесных деревьев.
Ранее было доказано, что волновое возмущение является обязательным атрибутом в поведении популяции лесных деревьев.
Тогда получается, что полная симметрия между тремя видами пробных площадок (кольцевые, прямоугольные и секторные) будет соблюдаться при квадратных пробных площадках размерами 5x5, 10x10, 15x15 и 20x20 м. Это нужно учитывать в экспериментах.
Уменьшение размеров центральной площадки до размера боковых пробных площадок позволит точнее вывить и сдвиг начала координат вторых составляющих модели, то есть начал колебательных возмущений по радиальным геодезическим направлениям. Пока они находятся в центральном квадрате материнской ели 20x 20 м.
Уменьшение пробной площадки вокруг центральной материнской ели, по-видимому, сузит границы места её произрастания.
Пространственный график распределения лесных деревьев (рис. 6.36) на выделе 8 кв. 37 существенно отличается от рис. 6.7 с горбами для отдельных биогрупп деревьев.
|
Рис. 6.36. Изменение плотности всех деревьев на пробной площади от расстояния и азимута на выделе 8 лесного квартала 37 |
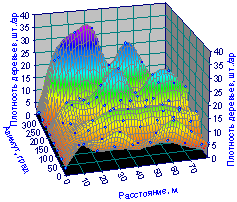
Здесь явно заметны не отдельные группы деревьев, а целые их «гряды», каждый из которых содержит возвышенности по радиальным направлениям.
Тогда понятно, что биоэнергетическое воздействие центральной ели на выделе 8 кв. 37 гораздо сильнее (активнее) по сравнению с материнской елью на выделе 9 кв. 30.
Заметно, что максимальные возвышенности вдоль окружностей кольцевых пробных площадок, то есть по азимуту, расположены в правильном геометрическом порядке.
Поэтому может оказаться, что на лучших по экологическим свойствам древостоях (а значит, и по техническим свойствам древесины) частично обособленные биогруппы лесных деревьев образуют своеобразную территориальную сетку.
Эта системная по фитоценозам сетка может иметь сильную территориальную и компонентную диффузию по площади и по видовому составу лесных деревьев и других видов растений, что требует новых экспериментов для доказательства процессов межвидовой диффузии.