
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

6.6. Плотность ели на пробной площади кв. 37
Исходные данные приведены в табл. 6.6.
Плотность деревьев ели в центре круговой площади очень низка и равна всего 0,95 шт./ар, хотя размеры имеет 20 x 20 м.
|
Таблица 6.6 Плотность деревьев ели nE, шт./ар
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
На четырех кольцевых пробных площадках наблюдается максимальное значение плотности деревьев ели. Средневзвешенная средняя плотность на круговой пробной площади равна 4,34 шт./ар.
Север. Малый доверительный интервал значений показателя по табл. 6.6 на прямоугольных площадках за центральным квадратом показывает на северном направлении (рис. 6.37) спокойное изменение основной тенденции по формуле
![]() . (6.35)
. (6.35)
|
Рис. 6.37. Тренд изменения плотности деревьев ели на пробной площадке в зависимости от расстояния в северном направлении |
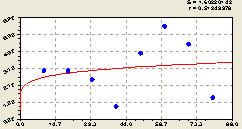
Здесь биотехнический закон превратился в закон показательного роста. Такое упрощение закономерности, как правило, показывает упрощенное поведение биологической популяции без экспоненциального торможения процесса роста по показательному закону.
Но при этом, как видно из расположения точек относительно графика на рис. 6.37, заметно сильное колебательное воздействие материнской ели на сородичей.
По остаткам (рис. 6.38) получилось более спокойное по сравнению со всеми деревьями уравнение
![]()
![]() . (6.36)
. (6.36)
Это спокойствие у деревьев ели выражается в том, что начальный период колебания равен 2 x 21,56103 ≈ 43,12 м.
|
Рис. 6.38. График волны изменения плотности деревьев ели по остаткам на пробной площадке от расстояния в северном направлении |
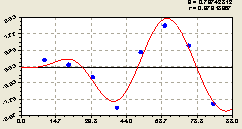
При этом начало волны колебательного возмущения находится на расстоянии 1,05486 x 21,56103 / π = 7,24 м в северную же сторону, то есть в сторону биоэнергетического воздействия ели на свои отпрыски.
Вместе (рис. 6.39) обе составляющие дали формулу конструкции вида
![]()
![]()
![]() , (6.37)
, (6.37)
в которой после утряски значения параметров модели изменились.
|
Рис. 6.39. График изменения плотности деревьев ели на пробной площадке в зависимости от расстояния в северном направлении |
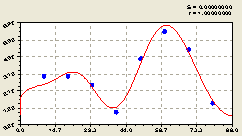
Так, например, начальный период успокаивающегося колебания стал равным примерно 33,48 м, а начало действия волны после уточнения находится на близком от центра расстоянии 0,089046 x 16,73948 / π = 0,47 м в северную же сторону. Этот эксцентриситет биоэнергетического воздействия уже сравним не только с диаметром надземной кроны, но и с диаметром ствола.
Восток (рис. 6.40) по радиусу круговой пробной площади или же по стороне света крестообразной пробной площади характеризуется биотехнической закономерностью
![]() . (6.38)
. (6.38)
Как всегда, то есть, как и в предыдущих статистических моделях данного типа, постоянный член показывает влияние местопроизрастания на генерацию подроста ели, а переменный член готовой статистической модели (6.38) показывает изменение биоэнергетического влияния центральной материнской ели в зависимости от расстояния до других деревьев ели от неё.
|
Рис. 6.40. Тренд изменения плотности деревьев ели на пробной площадке в зависимости от расстояния в восточном направлении |
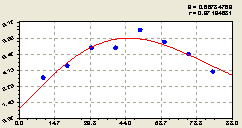
Здесь, по расположению точек относительно графика, видно, что через прямой угол по азимуту от северного направления волновое возмущении ослабло. Причем снижение значительное, и оно проявляется в основном на промежутке расстояния 30...50 м.
По остаткам (рис. 6.41) получена биотехническая закономерность
![]()
![]() . (6.39)
. (6.39)
|
Рис. 6.41. График волны изменения плотности деревьев ели по остаткам на пробной площадке от расстояния в восточном направлении |
Чем объяснить такое изменение по окружности? Пока нам неизвестно, и это становится реальным направлением тематики темой НИРС, то есть научно-исследовательской работы студентов - будущих бакалавров и магистров направлений подготовки «Защита окружающей среды» и «Природообустройство».
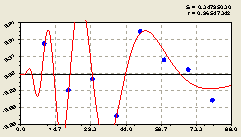
Вместе (рис. 6.42), после повторной параметрической идентификации всех полученных составляющих, получили уточненную формулу
![]()
![]()
![]() . (6.40)
. (6.40)
Читатель может спросить, а действительно ли формула (6.40) показывает влияние материнской ели? Может быть, при одном и том же возрасте нескольких деревьев ели, они совместно создают общее биополе и оказывают им на проростки общего молодого подроста? Четкого разделения объекта исследования у нас пока нет. Нужны измерения возраста деревьев. Тогда появится практическая возможность выявления динамического равновесия на территории и в компонентах древостоя и леса.
|
Рис. 6.42. График изменения плотности деревьев ели на пробной площадке в зависимости от расстояния в восточном направлении |
Из графика на рис. 6.42 видно, что компьютер полагает наличие сильной флюктуации на участке 10...30 м. но при этом частота колебания существенно снижается из-за того, что период возмущения быстро нарастает с удалением от материнского дерева ели.
Начиная от расстояния 50 м происходит плавное, но сильное, снижение биоэнергетического влияния центральной ели.
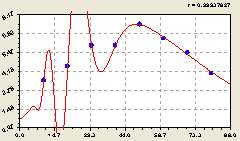
Юг. В этом направлении (рис. 6.43) получилась закономерность
![]() . (6.41)
. (6.41)
|
Рис. 6.43. Тренд изменения плотности деревьев ели на пробной площадке в зависимости от расстояния в южном направлении |
Заметно по расположению точек вокруг графика на рис. 6.43, что возможны даже две волны возмущения плотности у деревьев ели на пробных площадках на южном направлении крестообразной пробной площади на выделе 8 квартала 37.
По остаткам (рис. 6.44) получилось уравнение
![]() . (6.42)
. (6.42)
Оно показывает, что амплитуда изменяется по закону показательного роста, а частота биоэнергетического колебания постоянная. При этом отсутствует сдвиг начала волны, и поэтому начало координат находится на продольной вертикальной оси ствола центральной материнской ели. А какую форму (точечную, линейную, объемную) имеет начало волны? Об этом пока не можем даже догадываться. Но есть гипотеза, что учет формы и размеров кроны материнского дерева, а также формы и размеров её ствола, позволят подойти к моделированию. А полученные биотехнические закономерности дадут апостериорную информацию и идеи для выяснения теоретических подходов, в частности, к решению задачи взаимодействий деревьев в иерархии развития и роста.
|
Рис. 6.44. График волны изменения плотности деревьев ели по остаткам на пробной площадке от расстояния в южном направлении |
Вторая волна (рис. 6.45) получает амплитуду по полной конструкции биотехнического закона проф. П.М. Мазуркина. При этом половина периода колебательного возмущения убывает по закону Никитина как разность от начального периода около 30,89 м по формуле
![]()
![]() . (6.43)
. (6.43)
На других сторонах света позитивно адаптивной волны нет.
|
Рис. 6.45. График второй волны плотности ели по остаткам на пробной площадке от расстояния в южном направлении |
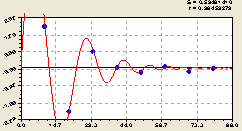
Таким образом, на южном направлении проявились два различающихся процесса. Что конкретно они показывают, этого мы пока точно не знаем.
По всей вероятности, может оказаться, что формула (6.43) в большей мере зависит от освещенности под пологом кроны самой материнской ели. А формула (6.42) кризисного роста возмущения по мере удаления от центральной ели происходит из-за ослабления её биополя, если предположить, что она растет одна, со своими отпрысками, и вокруг нет других материнских деревьев ели. Во всяком случае, эта гипотеза подходит для объяснения поведения деревьев по всем геодезическим направлениям.
Вместе (рис. 6.46) была получена закономерность
![]()
![]()
![]()
![]() . (6.44)
. (6.44)
Общая модель (6.44) показывает, что ближняя к центру ели волна является позитивной, направленной на адаптацию деревьев ели на круговой площадке радиусом почти до 50 м.
|
Рис. 6.46. График изменения плотности деревьев ели на пробной площадке в зависимости от расстояния в южном направлении |
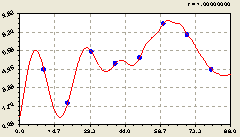
Дальняя волна с нарастающей амплитудой является кризисной, то есть характеризует кризисную адаптацию деревьев ели к условиям своего произрастания. Южное направление получило два колебательных возмущения среди плотности деревьев ели.
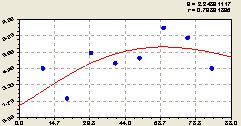
Запад. В эту сторону света распределение деревьев ели (рис. 6.47) снова стабилизировалась по закономерности
![]() . (6.45)
. (6.45)
|
Рис. 6.47. Тренд изменения плотности деревьев ели на пробной площадке в зависимости от расстояния в западном направлении |
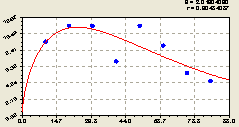
График на рис. 6.47 показывает классический пример поведения во многих явлениях и процессах природно-антропогенного или иного взаимодействия. Например, аналогично ведут себя виды и разновидности загрязнения воздуха в зависимости от расстояния от края автомобильной дороги.
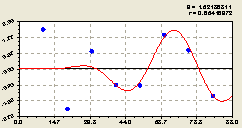
По остаткам (рис. 6.48) получена кризисная закономерность
![]()
![]() . (6.46)
. (6.46)
|
Рис. 6.48. График волны изменения плотности деревьев ели по остаткам на пробной площадке от расстояния в западном направлении |
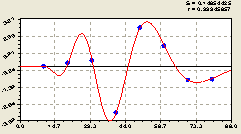
Иначе говоря, распространение от центра с материнской елью других сородичей происходит по классическому принципу распределения в зависимости от удаления от источника.
Плотность семян ели изменяется по такому же графику. Но большинство погибло из-за отсутствия условий для появления из семян всходов ели.
Вместе (рис. 6.49) после совместной параметрической идентификации трех составляющих получилась модель вида
![]()
![]()
![]() . (6.47)
. (6.47)
Через многие годы, с начала репродуктивного периода ели, образовалась сложная фигура плотности распределения деревьев ели.
|
Рис. 6.49. График изменения плотности деревьев ели на пробной площадке в зависимости от расстояния в западном направлении |
Однако, этому процессу возможно только одно объяснение как феномену, так как уже растущие ели мешали бы правильному распределению семян ветром от материнской ели.
Поэтому правильное теоретическое объяснение сводится к тому, что внутри древостоя происходит постоянно многолетнее влияние биополя материнской ели, активно и постоянно влияющей на проращивание семян именно в том месте, на котором нужно и на котором возможно по условиям леса. Однако само многолетнее влияние также имеет цикличный характер, - ведь максимальное плодородие ели наступает один раз примерно в семь лет.
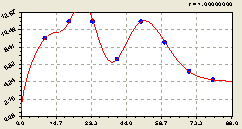
Всего на кольцах. Вышеизложенный процесс доказывается распределением (рис. 6.50) растущих деревьев ели по формуле тренда
![]() . (6.48)
. (6.48)
|
Рис. 6.50. Тренд изменения плотности деревьев ели на кольцевой пробной площадке от расстояния |
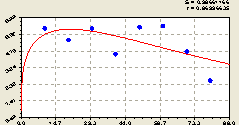
Активность биоэнергетического воздействия материнской ели на развитие и рост своих разновозрастных детей в среднем максимально проявляется на расстоянии 10...30 м.
По остаткам на рис. 6.51 получен график в соответствии с формулой
![]()
![]() . (6.49)
. (6.49)
|
Рис. 6.51. Плотность деревьев ели по остаткам на пробной площадке от расстояния |
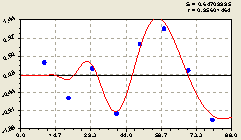
Этот интервал максимума можно измерять высотой материнского дерева в 23 м. Тогда получается, что высокая интенсивность биополя у любого дерева репродукционного возраста находится на интервале расстоянии от 0,5 H до 1,5 H.
Формула (6.49), показывающая кризисное изменение зоны влияния биополя от материнской ели по сложной амплитудно-частотной характеристике, в отличие от южного направления содержит торможение значений амплитуды. За пределами расстояния 80 м начинается её спад.
Вместе (рис. 6.52) постоянный член, стрессовое возбуждение и волновое изменение образуют после идентификации общую формулу
![]()
![]()
![]() . (6.50)
. (6.50)
Распределение плотности ели таково, что уже с края надземной кроны возникает максимум энергетического воздействия материнской ели на сородичей.
Затем по тренду (6.48) энергия воздействия убывает, вначале медленно, а затем ускоряясь к краю круговой пробной площади.
|
Рис. 6.52. График изменения плотности деревьев ели на кольцевой пробной площадке от расстояния |
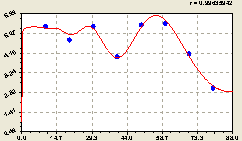
В центральном круге радиусом менее располагается само материнское дерево, то есть его ствол и наземная крона. Подземные кроны лесных деревьев живут по-особенному, отличаясь от поведения надземной кроны. Они выделяют только поросль. Но это уже совершенно иная физическая субстанция по сравнению с семенами. Поэтому по формуле (6.48) влияние на самого себя снижается до плотности 0,90505 шт./ар, то есть до уровня фоновой активности всего лесного участка на данной круговой площади. Корневая система при этом достигает, в пределе, троекратного роста в диаметре по сравнению с диаметром наземной кроны. В связи с этим, по-видимому, биоэнергетика корневых взаимодействий у лесных деревьев совершенно иная.
Пространственное распределение плотности деревьев ели (рис. 6.53)
|
Рис. 6.53 Изменение плотности деревьев ели на пробной площади от расстояния и азимута |
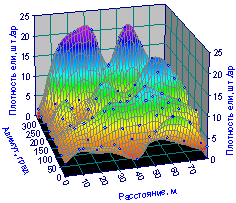
Сравнение с рис. 6.36 дает повод сделать вывод о том, что основу лесного древостоя дает распределение деревьев ели, а остальные породы выполняют соподчиненную роль.
Это видно из того, что:
во-первых, картины распределения схожи;
во-вторых, пространственное распределение ели по геодезическим направлениям с учетом сторон света и секторов между ними, проявляется четче по сравнению с распределением деревьев всех пород.