
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

6.7. Плотность подроста ели на пробной площади кв. 37
Как видно из данных табл. 6.7, имеется разница в плотности подроста ели по разным сторонам света. Существует также разница между сторонами света и кольцевыми пробными площадками.
|
Таблица 6.7
Плотность подроста ели
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Из-за разницы в площади территории, плотность подроста ели на центральном круге больше по сравнению с центральным квадратом на 100 (0,64-0,50) / 0,50 = 28,0 %.
В среднем по радиальным направлениям прямоугольных пробных площадок по убыванию плотности подроста ели все стороны света ранжируются по следующему предпорядку предпочтительности: запад (4,9 шт./ар), юг (3,5 шт./ар), восток (1,85 шт./ар) и север (0,55 шт./ар).
Север. Здесь (рис. 6.54), как и по всем параметрам в любой пространственной ориентации, соблюдается одна и та же закономерность
![]() . (6.51)
. (6.51)
|
Рис. 6.54. Тренд изменения плотности малых деревьев ели на пробной площадке в зависимости от расстояния в северном направлении |
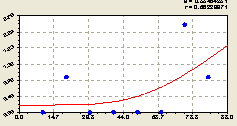
Однако тренд изменения плотности подроста ели, в зависимости от изменения радиуса от центральной материнской ели, имеет сложный характер.
Во-первых, имеются два пика импульсного изменения, которые можно идентифицировать двумя биотехническими законами.
Во-вторых, на оси абсцисс расположились пять точек с нулевой плотностью подроста ели. Ранее мы отмечали особую роль цифры «ноль» в лесной среде, то есть внутри лесной экосистемы.
По остаткам (рис. 6.55) была получена закономерность
![]()
![]() . (6.52)
. (6.52)
|
Рис. 6.55. График волны изменения плотности малых деревьев ели на пробной площадке по остаткам от расстояния в северном направлении |
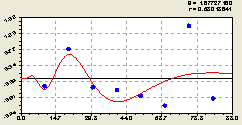
Нулевая численность подроста показывает, что лесная среда переполнена взрослыми особями деревьев. Поэтому остатки от тренда (6.51) по формуле (6.52) показывают отрицательную плотность подроста ели в соответствующих точках радиуса от центральной материнской ели.
Отрицательное значение показателя, как общеизвестно, показывает долг. Иначе говоря, лесная среда должна, хотя бы в будущем развитии и росте популяции лесных деревьев, сформировать из находящихся в лесной почве семян новые члены древесного сообщества.
Частично долг в подросте ели снимается второй волной по остаткам (рис. 6.56) в виде нарастающего по амплитуде колебания
![]()
![]() . (6.53)
. (6.53)
|
Рис. 6.56. График второй волны изменения плотности подроста ели на пробной площадке от расстояния в северном направлении |
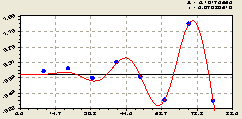
Двух волновое дополнение по плотности распределения ели было с южной стороны, а по подросту ели наблюдается в северном направлении. Конечно же, причина здесь не в солнечном освещении.
Тогда позитивное возмущение подроста ели на северной стороне света можно объяснить только антропогенным влиянием, да еще и со стороны лесной дороги, расположенной недалеко от материнской ели.
Вместе (рис. 6.57) четыре составляющие, вначале идентифицированные последовательно, после параллельной «встряски» имеют вид:
![]() , (6.54)
, (6.54)
![]() ,
,![]() ,
, ![]() ,
,
![]() ,
, ![]() ,
,
![]() ,
, ![]() , .
, .
|
Рис. 6.57. График изменения плотности малых деревьев ели на пробной площадке в зависимости от расстояния в северном направлении |
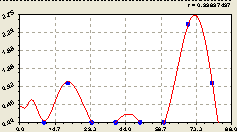
Модель (6.54) получила правильно упорядоченную запись, которая удобна при большом количестве получаемых составляющих.
График с двумя разрывами по оси абсцисс на рис. 6.57 убедительно показывает, что радиальное распределение подроста ели, хотя и не расстраивалось человеком, но имеет некоторые пустоты на ленточном лесном участке. Они, по-видимому, заполнятся проростками ели в недалеком будущем.
Восток определяется (рис. 6.58) трендом в виде формулы
![]() , (6.55)
, (6.55)
|
Рис. 6.58. Тренд изменения плотности малых деревьев ели на пробной площадке в зависимости от расстояния в восточном направлении |
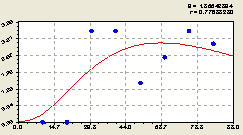
когда в расположении точек относительного него четко заметны два предельных кластера недалеко от материнской ели.
Первый кластер состоит из двух пробных площадок размерами 20 x 10 м на расстояниях по их центрам от материнской ели 10 и 20 м.
Здесь получилась нулевая плотность подроста ели. Второй кластер состоит также из двух пробных площадок и показывает достижение максимумов плотности у подроста ели на расстояниях их центров в 30 и 40 м. Такая кластеризация подроста ели не случайна. Она показывает сильную дифференциацию биогрупп ели по возрастному составу.
По остаткам (рис. 6.59) получилась закономерность вида
![]()
![]() . (6.56)
. (6.56)
|
Рис. 6.59. Волна плотности подроста ели на восток |
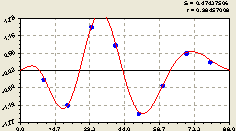
Трендовая зависимость (6.55) оставляет после себя остатки, по которым получилась высокоадекватная биотехническая закономерность. Коэффициент корреляции уравнения (6.56) равен 0,9847. Высокая адекватность была и в других примерах.
Вместе (рис. 6.60) составляющие дали готовую модель вида
![]() , (6.57)
, (6.57)
![]() ,
, ![]() ,
, ![]()
![]() ,
, ![]() , .
, .
|
Рис. 6.60. График изменения плотности подроста ели на пробной площадке от расстояния на восток |
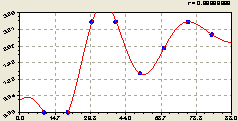
Высокая математическая адекватность модели (6.57), а также её отдельных составляющих при обособленном их моделировании, показывают высокую эвристическую ценность волновой теории размещения деревьев на выделе леса.
Юг (рис. 6.61) получает тренд, при каком-то красивом расположении точек, по формуле
![]() . (6.57)
. (6.57)
Здесь очень четко просматривается волна возмущения. Этот и предыдущие примеры высокоадекватного моделирования дают убедительную надежду на то, что со временем в лесной науке появится возможность сеточной теории лесного участка.
По остаткам (рис. 6.62) от формулы тренда (6.57) получили формулу
![]()
![]() . (6.58)
. (6.58)
|
Рис. 6.61. Плотность размещения подроста ели на юг в зависимости от расстояния |
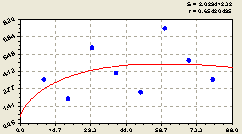
Сразу же видно, что эффект влияния солнечного освещения непосредственно около материнской ели здесь не просматривается. Виной этому является человек, многие годы заходивший с южной стороны со своими меркантильными интересами.
Поэтому вместе (рис. 6.63) три части дают:
![]() = n1 + n2 + n3 , (6.59)
= n1 + n2 + n3 , (6.59)
![]() ,
, ![]() ,
, ![]()
![]() ,
, ![]() .
.
|
Рис. 6.62. График волны изменения плотности малых деревьев ели на пробной площадке от расстояния в южном направлении |
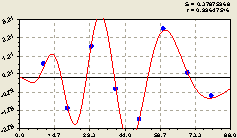
|
Рис. 6.63. График изменения плотности подроста ели на пробной площадке от расстояния в южном направлении |
Расположение точек на графике рис. 6.63 показывает, что пробные площадки по размерам в идеале должны соответствовать форме и размерам биогрупп лесных деревьев. Она даже простейшие формы при постоянных их размерах оказались эффективными, так как позволили получать высокоточные статистические модели, состоящие из суммы высокоадекватных биотехнических закономерностей.
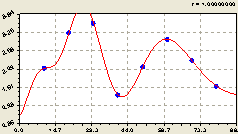
Запад. Мы теперь уже знаем конструкцию основной тенденции (рис. 6.64) и поэтому уверенно получили трендовую закономерность вида
![]() , (6.60)
, (6.60)
поэтому трудности в идентификации возникают только в первый раз.
|
Рис. 6.64. Тренд изменения плотности подроста ели на пробной площадке в зависимости от расстояния в западном направлении |
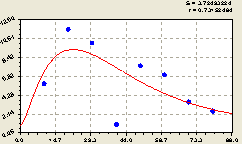
Знаем также, что вне зависимости от кажущейся хаотичности в расположении точек вокруг графика тренда, остатки будут моделироваться уравнением волновой функции.
На рис. 6.64 наблюдается провал плотности распределения подроста ели на расстоянии 40 м от материнской ели.
По остаткам (рис. 6.65) была идентифицирована закономерность
![]()
![]() . (6.61)
. (6.61)
|
Рис. 6.65. График волны изменения плотности малых деревьев ели на пробной площадке от расстояния в южном направлении |
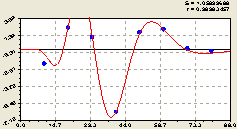
Получился почти симметричный график. Если у него площадь над осью абсцисс равна площади под координатной осью, то такая функция называется вейвлет-функцией.
Из-за применения биотехнического закона при описании параметров амплитуды и частоты колебания, вейвлет-функции распределения плотности ели становятся всегда асимметричными.
Вместе (рис. 6.66) постоянный член, стрессовое возбуждение и вейвлет-функция дают после повторной идентификации формулу
![]() , (6.62)
, (6.62)
n1 = 0,50000, ![]() ,
, ![]()
![]() ,
, ![]() .
.
График на рис. 6.66 показывает, что между точками в интервале расстояния 20...30 м может быть высокая плотность подроста ели.
|
Рис. 6.66. График изменения плотности подроста ели на пробной площадке в зависимости от расстояния в южном направлении |
В итоге получается что любая лесная среда пронизана потоками вещества и неизвестной по форме энергии, а также сигналов о свойствах и их отношениях с вещественно-энергетическими потоками.
Эти сигналы мы попытались расшифровать через размещение вещества деревьев на лесном участке, точнее его части в виде круговой пробной площади.
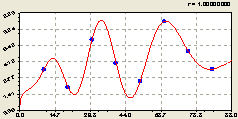
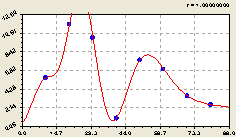
Всего по кольцам. По графику на рис. 6.67 получена модель вида
![]() . (6.63)
. (6.63)
|
Рис. 6.67. Тренд изменения плотности подроста ели на кольцевой пробной площадке в зависимости от расстояния |
Заметно, что общее радиальное распределение по концентрическим кольцевым пробным площадкам равносильно распределению на западном направлении.
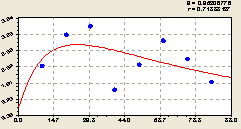
По остаткам (рис. 6.68) после формулы (6.63) была идентифицирована закономерность вида
![]()
![]() . (6.64)
. (6.64)
|
Рис. 6.68. Волна плотности подроста ели на юг |
Интегрированием уравнения (6.64) можно рассчитать площадь фигуры под графиком. Если она приближается к нулю, то формула (6.64) будет характеризоваться в полной мере как вейвлет-сигнал.
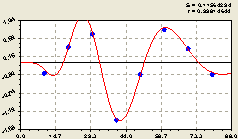
Вместе (рис. 6.69) образовалась закономерность
![]() = n1 + n2 + n3 , (6.65)
= n1 + n2 + n3 , (6.65)
![]() ,
, ![]() ,
, ![]()
![]() ,
, ![]()
|
|
|
Рис. 6.69. Плотность подроста ели на кольцевой пробной площади |
Асимметричные вейвлет-сигналы замечательны тем, что позволяют создать метод лесной инспекции по расхождениям фактических распределений плотности, а при подсчете ленточными таксационными способами и численности деревьев, с теоретически правильными вейвлет-сигналами.
На рис. 6.70 приведена пространственная картина распределения плотности подроста. Холмистая структура показывает куртинное распределение подроста ели.
|
Рис. 6.70. Изменение плотности подроста ели на пробной площади от расстояния и азимута |
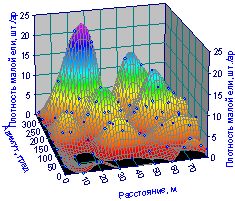
Разница между фактическим и теоретическим распределениями показателей древостоя на круговых пробных площадях и будет инт6гральным показателем качества структуры древостоя не только по отдельным лесным деревьям.
Этот показатель можно будет применять и по совокупностям лесных деревьев, их различающимся подгруппам, то есть кластерам в виде отдельных биогрупп, семейств биогрупп, куртин леса и пр.