
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

2. ВЗАИМООТНОШЕНИЯ НАСЕКОМЫХ С ЦВЕТКОВЫМИ РАСТЕНИЯМИ И ВНЕШНЕЙ СРЕДОЙ
В природе большинство видов насекомых с присущей им формой жизнедеятельности вступает во взаимодействие с растениями. Эти взаимоотношения касаются существенных сторон жизни видов и имеют значение для их выживания. Своеобразные взаимные отношения сложились между жалоносными перепончатокрылыми семейства пчелиных и цветковыми растениями (www.medovik.info).
Доступность широкого круга растений для пчелиных, благодаря их способности к быстрому перемещению в пространстве путем активного полета, богатый видовой состав, высокая численность популяций, биологические приспособления для сбора пыльцы и нектара, сделало их основными участниками процесса размножения цветковых растений. В то же время для пчелиных цветки растений являются основным источником пищи.
По литературным сведениям, пчелиные произошли от осоподобных предков вскоре после того, как начали появляться первые цветковые растения, которые развивались одновременно с пчелиными (www.medovik.info).
Перекрестное опыление, т. е. перенос пыльцы с цветка одного растения на цветок другого растения того же вида, в большей степени благоприятствует выживанию вида. Если бы у цветковых растений отсутствовали приспособления, обеспечивающие перекрестное опыление, то в большинстве случаев происходило бы самоопыление, так как собственная пыльца находится рядом с пестиком и вероятность ее попадания на рыльце большая. В результате биоразнообразие покрытосеменных растений, в силу незначительного набора пыльцы, было бы скудным.
Одно из приспособлений для предупреждения самоопыления у цветковых - пространственное разобщение пестиков и тычинок. Например, у двудомных растений мужские и женские цветки находятся на разных растениях, у однодомных пестики и тычинки развиваются в разных цветках того же растения, а у растений с обоеполыми цветками пространственное разобщение достигается в результате более тонких приспособлений (например за счет различия в длине столбиков и тычиночных нитей). Другой путь, препятствующий самоопылению, это разобщение во времени, т. е. неодновременное созревание пыльцы и рыльца пестиков (www.kedr.nago.ru).
В литературе высказываются предположения, что предки современных пчелиных различали цвета. Так А.А. Гроссгейм (1966) считает, что в ходе эволюции лепестки цветков приобрели заметную окраску. Это способствовало привлечению пчел. Поэтому яркая окраска цветков являлась условием, благоприятствующим перекрестному опылению, что сделало их более устойчивыми в борьбе за существование. Естественный отбор способствовал развитию различий между покрытосеменными по периоду цветения, окраске, форме и аромату цветков. Поскольку формы цветков становились более разнообразными, насекомые, посещавшие их, постепенно специализировались в собирании пищи с определенных форм цветков, оказываясь тем самым, менее приспособленными добывать ее с других форм цветков. Некоторые насекомые стали ассоциировать отдельные цвета и ароматы с наличием пищи и пренебрегать другой окраской и иными ароматами цветков. Между цветками некоторых растений и телом определенных видов насекомых установились соответствия, в результате чего они оказались полностью взаимозависимыми. Период цветения растений и время появления взрослых особей отдельных насекомых, посещающих цветки, также стали совпадать. Однако в целом у пчел не сложилось той тесной связи с определенными растениями, как у некоторых других насекомых, и они продолжали добывать пищу с растений широкого диапазона видов. Для растений было бы выгоднее, чтобы насекомые так сильно приспособились к их своеобразным цветкам, что были бы вынуждены ограничивать свои посещения узким кругом видов растений (www.medovik.info).
Медоносная пчела за свою жизнь посещает цветки разновидовых растений, но в каждый отдельный полет она может ограничивать свой выбор цветками растений одного вида. Однако при благоприятных условиях (например, высокое содержание сахаров в нектаре) пчела во время одного поискового полета может посещать цветки растений нескольких видов, что способствует их опылению. Кроме того, разнообразие пыльцы способствует развитию пчелосемей (П.М. Комаров, 1955).
Общеизвестно, что растения многих видов образуют пыльцу в количествах, превосходящих потребности воспроизведения. Большая часть пыльцы служит приманкой для привлечения насекомых, которые в значительной массе относят ее для питания в свои гнезда и только ничтожную часть ее переносят на рыльца других цветков. У некоторых растений сформировались приспособления, существенно уменьшающие количество пыльцы, которое могут собрать сборщицы нектара. Например, в цветках жабрея, тычинки расположены таким образом, что пыльца осыпается на ту часть спинки насекомого, с которой ему трудно ее счистить; причем в таком положении пыльцевые зерна приходят в непосредственное соприкосновение с рыльцами последующих цветков жабрея в тот момент, когда пчела проникает в них в поисках нектара. Тычинки портулака крупноцветного изгибаются в направлении прилетевшего на цветок насекомого. Если прикоснуться к тычинке иглой, тычиночная нить в течение нескольких секунд оклоняется, а затем постепенно возвращается в исходное положение. По прошествии нескольких часов после опыления цветки портулака закрываются. Цветки многих видов, например липы, привлекают пчел нектаром, который они выделяют в больших количествах (www.medovik.info).
Пыльцу пчелы одной семьи могут одновременно собирать с множества растений, но основная масса ее добывается с относительно небольшого числа видов, о чем свидетельствуют результаты наших исследований (Д.М. Панков, 2009). Из общего числа обножек, принесенных сборщицами пыльцы нескольких пчелиных семей, более 50% было собрано с бобовых, преимущественно с эспарцета песчаного и донника желтого, 15% - с деревьев и кустарников семейства розоцветных и около 10% - с ветроопыляемых растений (береза, вяз, тополь и др.). Таким образом, с этих растений пчелы экспериментальных семей собрали около 80% пыльцы.
Приемы сбора пыльцы насекомыми с цветков растений разных видов зависят от строения цветка. Собирая пыльцу с цветков розы и одуванчика, пчела быстро передвигается среди тычинок, часто пригибая их к себе и надкусив пыльники мандибулами, высвобождает пыльцевые зерна. С люпина пчелы собирают ту пыльцу, которой они обсыпаются во время поиска нектара; счищая ее с себя, они укладывают ее в пыльцевые корзиночки на задних ногах и несут в улей. Посещая цветки определенных видов, некоторые пчелы, счистив приставшую к ним пыльцу, затем отбрасывают ее в сторону, например, на подсолнечнике можно наблюдать, как одни виды пчел сбрасывают пыльцу, в то время как другие ее набирают (www.pcela.ru).
По мнению С.К. Кириленко (1987), посещать цветок пчелу побуждает окраска его венчика. Цветки на фоне зелени хорошо различаются пчелами-работницами благодаря более яркому (для их глаза) тону по сравнению с листьями (рис. 11).

Рис. 11. Цветок и листья лапчатки ползучей, сфотографированный через желтый (слева), синий (в середине) и ультрафиолетовый (справа) фильтры. Листья очень слабо отражают лучи во всех трех основных (для глаза пчелы) областях спектра. На всех фотографиях изображение листьев оказалось неотчетливым (по Даумеру).
На снимках лапчатки ползучей (рис. 11), сфотографированной через фильтры, соответствующие трем основным цветовым областям рабочих особей медоносной пчелы (желтая, синяя, ультрафиолетовая), листья в отличие от цветка, представляются значительно более темными.
Высокая чувствительность глаза пчелы-работницы к ультрафиолетовым лучам не только позволяет ей определять положение солнца в пасмурную погоду, но и способствует распознаванию цветов.
Сходные по форме и размеру цветки крестоцветных (желтушника, рапса и горчицы полевой) глазу человека представляются одинаково желтыми (рис. 12). Однако пчелы различают их хорошо. Когда же цветки покрывали стеклом, не пропускающим ультрафиолетовые лучи, пчелы не могли их различить. Для глаза человека становятся хорошо различимыми лишь снимки этих цветков, сфотографированных через ультрафиолетовый фильтр. Определено, что цветки горчицы отражают 9,5% ультрафиолетовых лучей, цветки рапса - 1,4%, а желтушник - практически не отражает таких лучей.
По мнению Даумера, пчела, приблизившись к цветку, исследует его дальше в том случае, если она способна уловить исходящий от него аромат. Подтверждение этому нашлось во время мечения пчел посещающих цветки. Если применявшаяся для мечения краска с резким запахом наносилась близко к голове пчелы, то ее поведение в течение нескольких минут становилось необычным. Когда она приближается к цветку, даже к тому, который часто посещала прежде, насекомое несколько минут летает перед ним, а затем опускается на лист или на другую удобную опору поблизости и начинает чистить антенны. Затем, снова приблизившись к цветку, она проникает в него или, немного полетав перед цветком с направленными вперед антеннами, опускается и опять чистит их. Если при мечении краска не попала на ее антенны, то спустя некоторое время она вскрывает цветок обычным образом. Вероятно, резкий запах растворителя в краске препятствует улавливанию пчелой значительно более слабого аромата цветков, к которым она приближается, а до тех пор, пока она не сможет воспринять его, т. е. пока растворитель не улетучится, она не опускается на цветок. Таким образом, если аромат цветков, посещаемых пчелами растений, замаскировать более сильным посторонним запахом, они неохотно вскрывают этот цветок до тех пор, пока присутствуют маскировочные запахи (http://www.medovik.info/).

Рис. 12. Цветы желтушника (а), рапса (б) и горчицы полевой (в), сфотографированные через желтый (слева) и ультрафиолетовый (справа) фильтры (по Даумеру)
Роль запаха цветка состоит в том, чтобы, во-первых, побудить энтомофила тщательно исследовать цветок, а во-вторых, служить опознавательным знаком для сборщиц нектара, которые уже ассоциируют его особенный запах с пищей. Однако сильный аромат цветков (например, от массы цветков боярышника или белого клевера) сам по себе достаточен, чтобы на расстоянии привлечь пчел к цветкам. Вместе с тем, они в состоянии находить объекты по зрительным признакам. Например, пчел можно приучить отыскивать лишенный запаха сахарный сироп на цветных карточках, также не издающих какой-либо запах; так же возможна их дрессировка на неароматные цветки (черника, брусника). Но пчелы запоминают запах легче, чем цвет. Если предложить им на выбор, с одной стороны, запах на который они были надрессированы, в тесной связи с новым для них цветом, а с другой - цвет, сочетавшийся раньше с сахарным сиропом, насыщенным незнакомым для них запахом, то пчелы практически всегда избирают первую пару, предпочитая запах, ассоциирующийся у них с кормом, и нейтральный цвет, сигнализирующему о пище цвету и постороннему - запаху (www.medovik.info).
Когда пчела опускается на цветок в поисках нектара, она исследует язычком в нем все узкие щели. Во многих цветках нектарники расположены именно в таких местах. Иногда их положение указывается цветными пятнами, которые человеческому глазу представляются контрастирующими с основным тоном цветка. При исследовании фотографическим методом, на цветках обнаружены узоры, скрытые от невооруженного глаза человека. Например, по рисунку, поглощающей ультрафиолетовые лучи части цветка, пять видов лапчатки (Potentilla) с одинаково желтыми цветками располагаются в последовательный ряд. В то время как у лапчатки прямостоячей ультрафиолетовые лучи поглощаются маленьким пятном в середине цветка, у ползучей область их поглощения несколько шире, у весенней она занимает ⅔ лепестка, у золотисто-желтой ультрафиолетовые лучи отражаются лишь краем венчика, образуя на нем кайму, а у кустарниковой они поглощаются всей поверхностью цветка (рис. 13). Однако ультрафиолетовый узор на цветках играет, по-видимому, большую роль не в распознавании пчелами цветков, а в отыскании ими нектарников или пыльцы уже после того, как они опустятся на цветок.
Так, у стальника (Ononis natrix) из сем. Мотыльковых, форма цветка приспособлена к посещению насекомыми. Лодочка, поглощая своей поверхностью ультрафиолетовые лучи, резко выделяется на фоне паруса (рис 14).

Рис. 13. Снимок в ультрафиолетовых лучах желтых цветков пяти видов лапчатки прямостоячей (а), ползучей (б), весенней (в), золотисто-желтой (г) и кустарниковой (д) (по Даумеру)
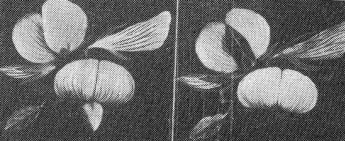
Рис. 14. Цветок стальника Ononis natrix, сфотографированный через желтый (слева) и ультрафиолетовый (справа) фильтры (по Даумеру).
Общая закономерность в распределении ультрафиолетового узора на цветках состоит в том, что центральная область цветка поглощает ультрафиолетовые лучи и их не отражает. Как только пчела переступит на лепестке невидимую для человека пограничную линию, отделяющую периферическую зону с ее ультрафиолетовым отражением от центральной, поглощающей эти лучи части, она легким толчком наклоняет голову и расправляет хоботок. Поэтому ультрафиолетовый узор следует рассматривать как указатель нектара (или пыльцы) для пчелы (www.fictionbook.ru).
В опытах с пчелами, никогда не посещавших цветков, обнаружен следующий факт: оказалось, что реакция наклона головы и вытягивания хоботка на ультрафиолетовый узор является врожденной.
Вход в более глубоко расположенные части цветка, где помещаются нектар и пыльца, выделяется не только окраской, но и своеобразным ароматом. У некоторых растений запах указателей нектара, оставаясь тем же по своему характеру, оказывается в них более сильным. Различия в запахе центральной и периферической частей венчика энтомофильных растений хорошо воспринимаются органами обоняния пчел, которые расположены на антеннах, в силу чего они могут быть приведены в тесное соприкосновение с любой поверхностью цветка, испускающей запах, в состоянии уловить эти различия, что облегчает пчеле поиск нектарников. Запаховые указатели нектара и пыльцы распространены среди цветков значительно шире, по сравнению со зрительными. Наряду с указателями цвета и запаха отысканию пчелой нектарников способствует особая трубчатая форма прилежащей к ним части венчика, воспринимаемая ее органами осязания.
Во время отбора нектара (рис. 15, 16) пчела, вероятно, воспринимает лишь качество пищи. Набрав корм, она перед возвращением в гнездо облетывается вокруг источника корма, знакомясь с его положением относительно ориентиров.
Цветки растений многих видов изобилуют нектаром и пыльцой лишь в определенное время дня. В остальные часы нектара в цветках бывает мало или значительно снижается концентрация сахара в нем, а пыльца в большинстве случаев вообще отсутствует. Благодаря хорошо развитому чувству времени пчелы приспосабливаются к подобной периодичности.
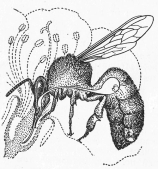
Рис. 15. Пчела, набирающая нектар в цветке. Показано положение в брюшке медового зобика с клапаном (по Реннеру)

Рис. 16. Пчела, набирающая сахарный сироп. Пахучий орган раскрыт, положение брюшка горизонтальное, крылья неподвижны (по Реннеру)
Взаимосвязь медоносной пчелы с цветковыми растениями на протяжении эволюции отразилась на составе секрета пахучей железы. Активной частью в нем служит гераниол - вещество, содержащееся во многих эфирных маслах растений. В секретах животных оно не встречается. У молодых пчел его нет, а у сторожевых (средний возраст 14 дней) содержится в среднем 0,3 мг. К тому времени, когда пчела становится сборщицей, его количество увеличивается втрое, достигая 1 мг, что свидетельствует о продолжающейся физиологической перестройке организма пчелы в конце ульевого периода (Бох и Шерер, 1963). Данная закономерность, вероятно, содействовала усовершенствованию связей между растениями и насекомыми, что выразилось в осуществлении своевременного опыления больших массивов растений. Преимущества опыления насекомыми не исчерпываются увеличением количества семян или плодов. Например, при обилии опылителей у некоторых бобовых повышается доля ранних цветков, что приводит к более раннему созреванию плодов и их однородности; у дыни, земляники - улучшается качество плодов. Поэтому медоносная пчела в настоящее время является важным опылителем энтомофильных культур.
На деятельность насекомых-опылителей, в т.ч. медоносных пчел существенное влияние оказывают факторы внешней среды. Из показателей, характеризующих освещенность, наибольшее экологическое значение имеют ее интенсивность, продолжительность и спектральный состав. Это зависит от времени дня и года, географического положения и рельефа местности, а также от состояния атмосферы (www.botsad.ru).
Характер освещения играет важную роль для пчел в связи со спецификой их светового восприятия, смещенного по сравнению с таковым у человека на ультрафиолетовую область. Пчелы воспринимают световые волны в диапазоне от 300 до 650 нм (от ультрафиолетового до оранжевого). Восприятие ультрафиолетового излучения дает возможность пчелам ориентировать по так называемому скрытому рисунку цветков.
Из общего потока солнечной радиации, поступающей на Землю, ультрафиолетовое излучение составляет от 1 до 5%, видимое (от фиолетового до красного) - от 15 до 45 и инфракрасное - от 49 до 84%. Интенсивность ультрафиолетового излучения резко уменьшается, когда угол на Солнце ниже 30° и возрастает при подъеме местности над уровнем моря.
В годовом цикле максимум солнечной радиации поступает на Землю во второй половине июня - начале июля. В безоблачные дни этого периода в равнинные районы умеренных и низких широт поступает 1,2-1,4 кал/см2мин солнечной энергии (Б.А. Айзенштат, 1981). При этом освещенность может достигать 100 тыс. лк. Облачность уменьшает освещенность до 20 тыс. лк. Минимальная освещенность приходится на декабрь. В безоблачную погоду этого времени года она достигает 10 тыс. лк, а при облачности - падает до 1000 лк. Интенсивность радиации уменьшается под покровом зеленых растений. Крона лиственных деревьев может задерживать до 98% солнечной радиации. Наряду с этим зеленые растения меняют спектр проходящего через них излучения (А.Н. Мартынов и др, 2008).
Изучение влияния освещенности на вылеты пчел из улья (Батлер и Фимпи, 1942) показало, что в течение одного дня число вылетающих из улья пчел возрастает при увеличении солнечной радиации на 0,1 калории на 1 см2/мин в среднем на 10-20%.
Таким образом, показатели, характеризующие освещенность, отличаются в зависимости от географии, местообитания, состава биоценоза, времени дня и года. Суточная и сезонная периодичность изменения освещенности спектрального состава света, а также связь этих изменений с динамикой общего биотического комплекса привели к развитию у насекомых фотопериодических адаптаций. Они выражаются в приспособленности жизненных циклов к определенной продолжительности дня. С этим, у многих видов насекомых, связана цикличность размножения, смена фаз индивидуального развития, активность репродуктивной функции, начало и окончание диапаузы (Практикум..., 2005).
По территории Бийской лесостепи простираются Колыванские увалы. Значительные площади сельскохозяйственных угодий СПОК «Возрождение 2», на землях которого проводились наши исследования, находятся у подножия увала, на его пологих склонах и на плоских широко простирающихся вершинах. Склоны, из-за крутизны, используются под сенокосы.
В ходе исследований было отмечено, что продуктивность пчелосемей, расположенных на возвышенности была значительно выше, по сравнению с пчелосемьями, расположенными у подножия увала. Контрольный привес улья, в ясную солнечную погоду, составил, соответственно - 6,4 и 4,2 кг/день.
Вылет пчел с ульев, расположенных на возвышенности, отмечен с 4 ч, интенсивная летная активность с 7 ч, в то время как у подножия увала, соответственно - 6 и 9 ч. По нашему мнению, это связано с тем, что на возвышенности увала, где отсутствуют густая древесная растительность, затенению ульи подвергаются меньше. Это способствует лучшей освещенности пчелосемей и активизирует их лет с ранних утренних часов, что положительно влияет на процесс опыления сельскохозяйственных культур.
По результатам двухфакторного дисперсионного анализа (Е.К. Еськов, 1990) суммарная сила влияния освещенности и трофического фактора сотавила 76±3%, из них 39±7% приходится на долю трофического фактора и 14±2% - на освещенность. Взаимное влияние факторов составляет 23±2%.
Однако, расположение пчелосемей на возвышенности увала не всегда позволяет получить высокую продуктивность улья и эффективное опыление. Нами установлено, что если пчелосемьи располагаются на возвышенности, а посевы энтомофильных сельскохозяйственных культур у подножия увала, пчелы не спускаются вниз, а посещают цветки только тех растений, которые выращиваются в пределах возвышенности.
Пчелосемьи, расположенные у подножия увала, в ясную солнечную погоду активно работают на травостоях в этой зоне, а так же наблюдается их активный лет на посевы, расположенные на возвышенности.
Такую разницу в поведении пчел мы объясняем тем, что пчела, возвращаясь в улей, несет в зобике до 60 мг нектара, а так же пыльцу, уложенную на задние конечности. Таким образом, масса груза может достигать 1/3 от массы пчелы, что создает дополнительные усилия и затраты энергии по доставке груза в улей, находящийся на возвышенности. Согласно наших подсчетов, проводимых в 2007 г., на посевах эспарцета у подножия увала и на возвышенности из расчета на 1м2 травостоя, цветки посетили в период активного лета в среднем, соответственно - 8-10 и 11-14 пчел/мин при наличии 6-8 пчелосемей на 1 га. На 4-5 день цветения травостоя эспарцет, возделываемый на возвышенности, был убран на сено. Однако количество пчел, посещающих растения эспарцета у подножия увала практически не изменилось. Пчелы, расположенные в ульях на возвышенности, посещали медоносы, удаленные на значительное расстояние от пасеки, хотя до травостоя эспарцета у подножия увала данное расстояние было в 2,5 - 3 раза короче.
Температура относится к числу показателей, лимитирующих летную деятельность пчел. В периоды похолоданий, когда температура опускается ниже 5°С, пчелы могут полностью прекращать вылеты.
Во время похолоданий, когда дневная температура не превышает 6 - 7°С, отдельные пчелы вылетают из улья. Слабый лет пчел отмечается при температуре 8 - 9°С. При 13 - 15°С происходит стабильный лет.
Жизнедеятельность пчелосемей во многом зависит от поддержания оптимальных условий в улье. Нектар, приносимый пчелами в улей, содержит 50 - 70% воды, тогда как в зрелом меде воды содержится не больше 20%. Следовательно, во время сильного взятка в улей ежедневно поступает значительное количество воды, которую нужно удалить из нектара при помощи испарения, осуществляемое за счет вентиляции. В жаркий день на прилетной доске можно видеть много пчел, которые быстрыми взмахами крылышек гонят из улья сильную струю воздуха. Этот воздух насыщен парами воды (medosbor.com/books/mal4.shtml).
Если улей имеет только один нижний леток и рамки сверху закрыты плотным положком, то на вентилирование пчелы тратят много энергии и это отвлекает их от работы на медосборе. Слабая вентиляция может вызвать духоту в улье, что часто приводит пчел в роевое состояние даже во время сильного взятка. Чтобы избежать указанных недостатков, нужно создать в улье условия для хорошей вентиляции. Для этого во время взятка во всех корпусах улья летки открывают полностью, а между корпусом улья и отъемным дном спереди вставляют клинышки, в результате чего образуется широкий проход (Е.К. Еськов, 1990).
Если в корпусах, поставленных на гнездо, нет летковых отверстий, то корпуса «расклинивают», то есть вставляют между ними клинышки, создавая щели для прохода воздуха. Сверху на рамках нужно оставить только холстик, завернув его передний край так, чтобы образовался проход шириной 4-5 см; если гнездо закрыто потолочными дощечками, то их надо раздвинуть. Между крышкой улья и верхним корпусом так же следует вложить клинышек для образования прохода. Это позволяет создать в улье тягу воздуха и семья легко справляется с удалением паров воды.
Резко снижается работа пчел на медосборе при сильном нагревании ульев солнцем. Поэтому ульи целесообразно размещать около деревьев, затеняющих их в жару. Если пасека стоит на открытом месте, то целесообразно во время сильного зноя накрывать ульи скошенной травой или ветвями деревьев.
Ветер оказывает существенное влияние на жизнедеятельность пчел. При ветреной погоде выдувается тепло через леток и стенки улья. Для поддержания тепла в таких условиях пчелы расходуют много корма и энергии. В результате семьи развиваются значительно медленнее, чем в местах, хорошо защищенных от ветра. Кроме того, ветер способствует налетам пчел одних семей на другие, затрудняет ориентировочные облеты молодых пчел и маток. В местах, где часто дуют ветры, пчелы становятся более раздражительными.
По мнению А. Нестерова (2010), сильный ветер весной снижает температуру окружающего воздуха и принуждает насекомых к бездеятельности. Гибель пчел в холодные весенние дни в одной семье может достигать 3 тыс. особей, что соответствует 3/5 всей пчелосемьи.
Средняя скорость полета пчел около 22 км в час, поэтому в ветреные дни пчелы летают на более короткие расстояния и возвращаются без обножки.
Скорость воздушного потока измеряли на высоте 1 и 5 м при температуре 15 - 20ºС, когда отмечается стабильный лет пчел. Скорость ветра на высоте 5 м почти вдвое выше, чем на уровне 1 м от поверхности земли. Согласно данных Е.К. Еськова (1990), увеличение скорости ветра до 12-15 м/с уменьшало летную активность более чем в 9 раз (табл. 5)
Таблица 5. Влияние скорости ветра на лет пчел (по Е.К. Еськову, 1990
|
Скорость воздушного потока на высоте 5 м от поверхности земли, м/с |
Количество пчел, вылетевших из улья за 1 мин., шт |
|
до 1 |
62-65 |
|
5-8 |
26-30 |
|
12-15 |
5-7 |
Таким образом, на летную деятельность пчел оказывает влияние даже небольшие скорости ветра. Поэтому пчелосемьи нужно предохранять от ветра. Положительный эффект оказывает размещение пчелосемей у лесополос, сооруженных поперек господствующего направления движения ветрового потока. Кроме того, в жаркие периоды в полдень, лесополосы создают тень, что препятствует перегреву пчелосемей. При смещении солнца в последующие часы, ульи вновь подвергаются солнечному воздействию, что положительно сказывается на их летной деятельности.
Количество водяных паров, содержащихся в воздухе, непостоянно и зависит от уровня увлажнения, температуры и атмосферного давления. Максимально возможное насыщение воздуха водяными парами при нормальном атмосферном давлении возрастает с повышением температуры: при температуре 1ºС для насыщения 1 м3 воздуха требуется 4,84 г воды, при 10 - 9,33, при 20 - 17,32, при 30 - 30,04 и при 39°С - 48,20 г. В состоянии насыщения при неизменной температуре и давлении водяные пары находятся в нейтральном равновесии с водяной поверхностью. Понижение или повышение температуры нарушает это равновесие: при понижении происходит конденсация части водяных паров, при повышении - дополнительное насыщение за счет испарения (www.uvlazhniteli.ru).
Влажность воздуха рассчитывается на основе измерений содержания в нем водяных паров. Для ее характеристики используют следующие показатели (Т.М. Савцова, 2008): 1) абсолютная влажность - количество водяного пара в единице объема (г/м3); 2) относительная влажность - отношение упругости водяного пара, содержащегося в воздухе, к максимальной упругости водяного пара, насыщающего пространство над плоской поверхностью воды при данной температуре (%), или отношение количества водяных паров в воздухе при данной температуре к тому количеству, которое требуется для полного насыщения воздуха без изменения температуры; 3) точка росы - температура, которую примет воздух, охлажденный при постоянном давлении, до состояния насыщения содержащихся в нем водяных паров; 4) парциальное давление (упругость) водяного пара - давление, которое приобрел бы пар, если бы занимал весь объем смеси при данной температуре (мм ртутного ст. или мб); 5) удельная влажность - количество водяного пара в 1 кг влажного воздуха (г/кг); 6) отношение смеси - количество водяного пара в 1 г сухого воздуха (г/кг); 7) дефицит влажности - разность между максимальной и фактической упругостью водяного пара при данной температуре и давлении.
Из перечисленных показателей, используемых для характеристики влажности воздуха в пчелином жилище в разные периоды годового цикла жизни семьи, наиболее полное представление о гигрорежиме дает относительная и отчасти, абсолютная влажность. Относительная влажность особенно важна тем, что физиологическое воздействие и восприятие гигрорецепторами насекомых равного количества водяных паров, содержащихся в воздухе, существенно изменяется от температуры.
Наблюдается зависимость между активностью работы медоносных пчел и наличием источников воды на пасеке. Нами отмечено, что при наличии поилок около пчелосемей, за 1 мин в ясную солнечную погоду в улей возвращается с обножкой 65-70 пчел, в то время как при отсутствии поилок данный показатель снижался до 50-55 шт/мин. При отсутствии поилок, на близлежащих водоемах наблюдалась массовая посещаемость пчел, тогда как при наличии поилок около ульев посещение водоемов пчелами было не значительным. Оборудование пасек поилками способствует активизации опылительной деятельности медоносных пчел за счет сокращения времени и энергии на поиск и транспортировку воды в улей.
Нами отмечено, что пчелы более охотно посещают поилки с подсоленной водой, чем с пресной. Данные показатели составили, соответственно - 40 и 160 шт/мин из расчета на 10 пчелосемей. Соль способствует нормализации минерального обмена в организме пчелы, что положительно сказывается на продуктивности улья в целом. При увеличении минерализации воды существенных отклонений в посещении поилок пчелами не наблюдалось.
Было установлено, что при наличии поилок только с пресной водой пчелы в поисках минерального питания часто посещают солонцы и др. источники минеральных солей. На поиск тратится значительное время, что отрицательно сказывается на эффективности пчелоопыления растений и медосборе. И наоборот, оборудование пасек поилками только с минерализованной водой, вынуждает медоносных пчел к поиску пресной воды. Поэтому наличие на пасеке поилок с пресной и минерализованной водой позволяет существенно активизировать работу пчел на цветущих массивах энтомофильных культур.
Геомагнитные и электрические поля входят в сложный комплекс геофизических факторов, различающихся между собой по характеру и результатам воздействия на биосферу. К строго векторизованным факторам относится гравитация. Гравитационная сила ослабляется обратно пропорционально квадрату расстояния от центра Земли. Поскольку гравитационное поле обладает всепроникающим действием, его невозможно ослабить посредством экранирования (Е.К. Еськов, 1990).
Специфическую структуру имеет магнитное поле Земли. На северном магнитном полюсе напряженность поля составляет около 0,6; на южном - 0,7; а на экваторе - 0,4 Э. Постепенный ход изменения напряженности от экватора к полюсам нарушается во многих географических зонах, образующих магнитные аномалии. Их различают на положительные и отрицательные. В районах положительных аномалий напряженность выше, а в отрицательных - ниже характерной для данной широты в других районах. Очень высокая положительная аномалия известна в Курской области, где напряженность магнитного поля достигает 1-2 Э, отрицательная - в южной оконечности Южной Америки (0,25 Э), а на территории России - в междуречье сибирских рек Ангары и Илима (Е.К. Еськов, 1990).
Небольшие изменения магнитного поля происходят в течение суток. Эти вариации (по горизонтальной составляющей до 20-30 гамм) связаны с токами в ионосфере, величина которых зависит от суточных колебаний ультрафиолетового излучения Солнца (www.krugosvet.ru).
Возмущения магнитного поля (бури), коррелирующие с солнечной активностью, имеют четко выраженный 27-дневный и 11-летний циклы (Е.К. Еськов, 1990).
Естественное распределение электрических и магнитных полей изменяется под влиянием антропогенного фактора. Искусственные поля многократно превосходят естественный фон. Например, в лабораториях создаются электромагнитные поля напряженностью до нескольких тысяч эрстед, а поле Земли составляет в среднем около 0,5 Э (MetaMir.wpdom.com).
Электрификация и расширение сети высоковольтных линий электропередачь существенно изменили характер распределения напряженности электрического поля.
Сеть высоковольтных линий электропередачь ежегодно возрастает. По многим из них передается переменный ток промышленной частоты (50 - 60 Гц). Напряженность электрического поля в зоне ЛЭП зависит от напряжения, передаваемого по линии. Максимальный уровень электрического поля вблизи ЛЭП 110 кВ составляет 33, ЛЭП 750 кВ - 155 В/см. Для многих наземных биологических объектов важна напряженность электрического поля у поверхности Земли. Она отличается у линий разных классов и зависит от высоты опор, провисания проводов и рельефа местности. Среднее значение напряженности электрического поля на высоте 2 м от земли под ЛЭП 500 кВ/см сотавляет 60 В/см, ЛЭП 750 кВ/см- 110 В/см, ЛЭП 1500 кВ/см - 174 В/см (Е.К. Еськов, 1990).
Эффективность действия низкочастотных электрических полей на пчел зависит от их численности. По данным акустического контроля семья, содержащая 1,8-2,2 тыс. особей, начинает возбуждаться под действием электрического поля промышленной частоты при напряженности 8,8+0,40 В/см (Qr = 19%). Семья с численностью 9-12 тыс. особей возбуждается при 5,2+0,41 В/см (Qr = 39%). Минимальная напряженность, необходимая для возбуждения слабой семьи, составляет 7 В/см, максимальная - 12 В/см; сильной - соответственно 3 и 9 В/см (Е.К. Еськов, 1990).
Чувствительность к току различных частей тела насекомого зависит как от их электропроводности и раздражимости, так и от многих случайных факторов, влияющих на величину контактного тока, например наличия в местах контактирования воды, частиц пыли, воска, прополиса и т. п. По этой причине порог чувствительности варьирует в широких пределах. Однако его величина имеет четкую тенденцию к понижению с увеличением численности особей в гнезде, т.к. с этим связано повышение частоты их контактирования.
Влияние электрического поля ЛЭП на эффективность летной деятельности пчел прослежено Е.К. Еськовым (1990) на пчелосемьях, находившихся под ЛЭП и на расстоянии 10 и 50 м от линии. При этом металлические крышки у части ульев, расположенных под ЛЭП, были заземлены. Это снижало напряженность электрического поля до 11 В/см (у незаземленных она составляла 75 В/см).
Оказалось, что в то время, когда растения слабо или совсем не выделяли нектар, масса содержимого медового зобика была наибольшей у пчел, вылетавших из незащищенных ульев, расположенных под ЛЭП - 2,3±0,08 мг и на расстоянии 10 м от нее - 2,0±0,06 мг. В это же время в зобиках пчел, подвергавшихся меньшему влиянию электрического поля ЛЭП, содержался меньший запас корма: у пчел, вылетавших из ульев под ЛЭП с заземленной крышкой, масса содержимого зобика составляла 1,4+0,04 мг, а из удаленных на 50 м - 1,5±0,03 мг. В период обильного нектаровыделения масса содержимого медовых зобиков у вылетающих пчел возросла во всех сравниваемых группах. При этом она осталась наименьшей у пчел, живших в защищенных ульях (2,1±0,22 мг) и в 50 м от линии (2,2±0,07 мг). У пчел из незащищенных ульев, размещенных под ЛЭП, в зобиках находилось 2,7±0,10 мг корма, а в 10 м от линии - 3,7±0,23 мг.
В условиях, когда медосбору препятствуют неблагоприятные условия, пчелы доставляли наименьшее количество нектара в незащищенные ульи, находившиеся под ЛЭП (по 9,4±0,45 мг) и в 10 м от линии (9,1±0,54 мг). В это же время пчелы, жившие в защищенных ульях под ЛЭП и в 50 м от нее, приносили соответственно по 12,8±0,84 и 11,2±0,75 мг нектара. Различия по массе содержимого медовых зобиков возросли в период интенсивного нектаровыделения: под ЛЭП без защиты 19,6±0,79 мг, с защитой 28,7±1,33, в 10 м от ЛЭП 21,5±1,2 и в 50 м от ЛЭП 28,8±1,46 мг (Е.К. Еськов, 1990).
Наименьшей продуктивностью отличались семьи, которые находились под ЛЭП. Они собрали в среднем по 31,6±2,90 кг меда. Немного больше его было собрано семьями, размещаемыми в 10 м от ЛЭП - 38,4±4,40 кг. Наиболее продуктивными оказались семьи, расположенные в 50 м от линии. Их медосбор составил 49,6±3,52 кг, что в 1,6 раза превосходило по медопродуктивности семьи, находившиеся без защиты под ЛЭП. Действие защиты позволило семьям интенсивнее пополнять кормовые запасы. Они собрали по 46,3±4,05 кг меда, максимально приблизившись по продуктивности к семьям, расположенным в 50 м от ЛЭП (Е.К. Еськов, 1990).
Понижение медопродуктивности пчелиных семей под действием электрического поля ЛЭП можно объяснить уменьшением воспроизводства рабочих особей и снижением продуктивности летной деятельности. Активность лета пчел не имела достоверных расхождений в семьях, подвергавшихся различному влиянию электрического поля ЛЭП: в ульи, находившиеся в 50 м от линии, возвращалось в течение минуты 127 особей, ЛЭП без защиты - 131, ЛЭП с защитой - 119 пчел. Высокий коэффициент вариации количества пчел, возвращавшихся в незащищенные ульи под ЛЭП, отражает резкие колебания летной активности. Это выражалось в кратковременных спонтанных резких увеличениях численности вылетающих пчел, которые полетав вблизи улья, вскоре возвращались в него (Е.К. Еськов, 1990).
Пчелосемьи расположенные у ЛЭП отличаются большей агрессивностью, чем семьи удаленные от ЛЭП. Она сохранялась на протяжении всего периода нахождения семей под линией. У летков отмечалось большое количество пчел. Для них характерен высокий уровень двигательной активности в пределах улья (Д.М. Панков и др, 2007).
Особенность поведения пчел под ЛЭП заключается в стремлении заделать воском и прополисом не только места соединения частей улья, но и летковые отверстия.
Несмотря на отрицательное отношение пчел к электрическому полю ЛЭП, не обнаружено их стремления покинуть эту зону. Не отмечено повышения ройливости пчел, находящихся под ЛЭП (Е.К. Еськов, 1990).
Расширение сети радио- и телевизионной связи, а также систем радиолокации сопряжено со значительным насыщением атмосферы электромагнитными полями. Для радиотелефонной связи используются сверхдлинные радиоволны частотой 3·10-3 - 3·10-2 мГц, в радиосвязи и телевидении - от длинных (3·10-2-3·10-1) до ультракоротких (до 3·104) и в радиолокации - от 3·104 до 3·105 мГц. Волны указанных частот в обычных условиях содержания пчел, по-видимому, не влияют на их состояние и жизнеспособность. Так, при воздействии на пчел полями, используемыми в телевидении и радиолокации, с плотностью потока в несколько десятков мВ/см2 не выявлено каких-либо специфических форм реагирования, изменения состояния или снижения жизнеспособности (Е.К. Еськов, 1990).
Для выявления устойчивости пчел к ультравысокочастотному облучению Е.К. Еськов (1990) использовал УВЧ генератор (частота около 40 мГц). Пчелы погибали в электрическом поле УВЧ через 6,1 + 1,01 мин, Qr= 133% (минимум - через 1,5, максимум - через 39 мин).
Перед грозой нередко можно наблюдать, как большое количество пчел без груза возвращается в улей. Летная деятельность прекращается, хотя освещенность, температура и сила ветра находятся в пределах оптимума. Наряду с этим, иногда перед грозой, у пчел повышается агрессивность по отношению к особям, подлетающим к летку и людям, находящимся на пасеке.
Одна из причин, влияющих на поведение пчел при приближении грозы - изменение напряженности электрического поля. Его порождают грозовые разряды. Регистрируя изменение электрического состояния атмосферы и наблюдая за поведением пчел. У. Варнке (1973) обнаружил наличие между этими процессами определенной связи. Он заметил, что агрессивность пчел достигает максимума с приближением грозового фронта до 600-800 км, когда скачкообразные (на 100 В/см и выше) изменения напряженности электрического поля на частоте 10 кГц повторяются через 1,2-0,5 мин.
Возвращение пчел в улей перед грозой и повышение агрессивности с приближением грозового фронта и активизацией атмосферы объясняются ростом напряженности и вариабельности электрического поля. Наводимые ими токи раздражают пчел при контакте с токопроводящими поверхностями, что вынуждает пчел возвращаться в гнездо. Вероятно, эта реакция входит у пчел в наследственно закрепленный адаптивный комплекс, обеспечивающий уменьшение гибели при неблагоприятных погодных условиях.
Агрессивность пчел друг к другу и людям, находящимся вблизи ульев, при приближении грозового фронта можно объяснить двумя причинами. Первая из них связана с возбуждением пчел, вынужденных возвращаться в улей без нектара и пыльцы. Это в некоторой степени сходно с поведением пчел в то время, когда они из-за отсутствия цветущих медоносов не могут пополнять свои кормовые запасы. В это время их агрессивность возрастает. Другая причина связана с тем, что прикосновение к человеку или насекомому вызывает раздражение наведенным током, протекающим в местах контактирования. Эта ситуация сходна с той, которая используется для получения яда посредством раздражения пчел током. Таким образом, известные аномалии поведения при приближении грозы связаны в основном с изменением электрического поля (Е.К. Еськов, 1990).
Наши исследования о влиянии электрического поля на летную активность пчел показали, что при медосборе на посевах эспарцета контрольный привес улья, находившегося в радиусе 10-12 м от ЛЭП 500 кВ, составлял 2,1-2,3 кг/сут. Суточный привес улья, находившегося на расстоянии 60-70 м от источника электрического поля достиг 3,5-3,8 кг/сут.
Количество пчел, возвращавшихся в улей с обножкой, расположенный в радиусе 10-15 м от источника электрического поля в период активного лета, составляло 60-65 особей/мин, в то время как в улье, расположенном в радиусе 60-70 м от ЛЭП 500 кВ, данный показатель увеличился до 80-85 особей/мин.
Таким образом, улучшить показатели работы медоносных пчел на посевах эспарцета и других сельскохозяйственных культур можно при размещении пасеки, где воздействие электрического поля будет минимальным. Это позволит увеличить продуктивность улья и получить высокие урожаи семян.