
Научная электронная библиотека
Монографии, изданные в издательстве Российской Академии Естествознания

1.1. Особенности северного биосырья с повышенным содержанием и более широким спектром разнообразия физиологически активных веществ (ФАВ)
До недавнего времени сырьевая база Якутии для фармацевтической промышленности использовалась очень незначительно. Возможно, этому способствовало бытовавшее теоретическое представление, согласно которому содержание ФАВ в растениях закономерно снижается с возрастанием Географической широты места произрастания [Макаров, 1989]. В свете этого представления, именуемого широтной теорией формирования физиолого-биохимических особенностей растений, поиски во флоре якутского региона, характеризующегося крайне суровыми природными условиями, сколько-нибудь действенных лекарственных растений представлялось заведомо бесперспективным делом.
Начиная с 40–50-х годов XX столетия, в Якутии начали проводиться широкие биохимические исследования с целью выявления региональных особенностей химического состава и питательной ценности местных кормовых растений. Наряду с основным, белково-углеводным обменом растений, изучались их микроэлементный и минеральный состав, вопросы физиологии, а также биосинтеза в растениях ФАВ – витаминов, алкалоидов, сапонинов, танинов и др. [Егоров и др., 1945; Егоров, 1954, 1969; Сергеев, 1954; Макаров, 1962, 1963, 1965, 1969, 1970, 1972, 1975, 1989; Самарин, 1962, 1965а, 1965б; Слепцова, 1971, 1972, 1977; Говоров, Торговкина, 1974; Габышев, 1977; Соболева и др., 1977; Попова, 2003]. Большой вклад в изучение роли климата на образование растениями различных химических веществ внес Н.И. Шарапов [1954, 1962]. В своих работах он показал, что влияние климата очень многосторонне и разнообразно. Климат в прошлом и теперь создает границы распространения видов растений, определяет состав растительности зон и областей, существенным образом влияет на всю жизнь растений, в том числе на способность создавать и накапливать важнейшие вещества. Им были сформулированы основные положения широтно-климатической (ландшафтно-климатической) теории биосинтеза веществ в растениях. Эта теория получила широкое распространение и признание. Согласно Н.И. Шарапову не все факторы внешней среды имеют одинаково важное значение для растений. Признавая исключительную роль фактора влаги, автор пришел к следующему заключению: оптимальные или умеренные температуры окружающей среды (воздуха, почвы) и полная обеспеченность влагой способствуют созданию и накоплению запасных безазотистых веществ в растениях, в частности сахаров, крахмала, жира или масла, клетчатки и пр. Наоборот, высокая температура окружающей среды, относительный недостаток влаги, сухость воздуха и почвы в течение вегетационного периода стимулируют образование и накопление белковых веществ (в том числе белков регуляторного действия) и менее благоприятны для синтеза углеводов.
В этом отношении Якутия является уникальным примером совместного действия климатических стресс-факторов на растительные и животные организмы. Наиболее яркими климатическими особенностями Якутии [Гаврилова, 1998] являются:
– Сочетание суровой продолжительной и малоснежной зимы с коротким, но относительно теплым и засушливым летом. По абсолютному минимуму (-60 °С) и годовой амплитуде температур (102 °С) Якутия не имеет аналогов в северном полушарии. Это связано с высокой прозрачностью атмосферы и малой облачностью, способствующих быстрому повышению или снижению температуры воздуха, а значит, возникновению частых осенних и весенних заморозков. Например, весна в Якутии на 2 недели короче, чем в Свердловской и Архангельской областях. Снег тает быстро, большая часть влаги испаряется (возгоняется) солнечным теплым днем или холодной ночью, за счет вымораживания при резком усилении ветра. Превышение температуры воздуха отметку +5 °С (вегетативная весна) устанавливается во второй половине мая и заканчивается в середине сентября. Весенние заморозки, которые сопровождаются интенсивным охлаждением почвы, бывают до первой половины июня, а с 15–20 августа отмечаются осенние. Продолжительность безморозного периода (Центральная Якутия) в среднем колеблется от 75 до 90 дней, а период благоприятный для вегетации растений составляет 60–80 дней [Гаврилова, 1998].
– По интенсивности солнечной радиации Якутск не уступает г. Ташкенту и Феодосии, то есть радиационный баланс в целом благоприятен для нормального развития растений. Однако, резкое нарастание солнечной радиации в начале вегетации растений (май), приводит к отрицательному эффекту за счет дисбаланса между необходимым для транспирации растений количеством воды и ее медленным поступлением к корням при пониженных температурах почвы. Продолжительность солнечного сияния в июне-июле не менее 18 часов в сутки. Средняя температура июля +18–190 °С (абсолютный максимум 38 °С), а января – 35–40 °С. Теплозащитная роль снегового покрова, несмотря на небольшую высоту, очень значительна, что имеет огромное значение для зимующих растений. Продолжительность залегания устойчивого снежного покрова составляет около 200 дней.
– Одним из главных лимитирующих факторов для произрастания растений является влага. По количеству осадков Якутия приближается к степным и полупустынным районам. Особенно засушливой бывает первая половина лета; основная часть осадков выпадает во второй половине лета и осенью. Осадков в г. Якутске за май – июнь выпадает 51 мм, а за июль-август – 85 мм [Гаврилова, 1998].
– К неблагоприятным климатическим условиям летом относятся суховеи, которые снижают тургор листьев, приводят к торможению ассимиляции и роста растений. Относительная влажность воздуха летом составляет в среднем 30–50 %. Это способствует интенсивному испарению влаги из верхнего почвенного слоя.
– Якутия расположена в области повсеместного распространения многолетнемерзлых грунтов. Они оказывают сильное влияние на почвенный климат, рост и развитие растений, создают очень своеобразную динамику частичной сезонной оттайки и водного режима.
В Центральной Якутии широко распространены подземные льды, глубина залегания верхней кровли которых варьирует от 1,5 до 3,5 м. Возраст многолетней мерзлоты Якутии исчисляется 600 тысячами лет. В засушливых условиях Якутии наличие такого мощного консерванта влаги может рассматриваться как положительный природный фактор. Однако низкие температуры грунтов, обусловленные многолетней мерзлотой (особенно в первой половине лета) отрицательно сказываются на интенсивности почвообразовательных процессов и продуктивности растений.
Таким образом, особые экологические факторы (температурно-влажностные, солнечно-инсоляционные, почвенные) Якутии инициировали в процессе эволюции живых организмов образование у них эффективных способов приспособлений, направленных на выживание в этих условиях.
Территория Якутии является своего рода резерватом генетического и ландшафтного разнообразия общемирового значения. В её флоре известны 88 видов трав, 26 кустарников и кустарничков, 7 деревьев – лекарственных растений, которые разрешены для использования в научной медицине и применяются в народной медицине как источники повышенного содержания и фракционного состава ФАВ и их структурного разнообразия [Голдовский, 1941; Егоров, 1945, 1954, 1969; Гриневич и др., 1977, Брехман, Гриневич, 1978; Телятьев, 1987; Брехман, Нестеренко, 1988; Алексеев и др., 1983; Макаров, 1989; Кершенгольц и др., 1995, 1996, 1999, 2002, 2004, 2005; Кузьмина, 2002]. Установлено, что по мере повышения степени экстремальности условий произрастания растений (обитания животных), например, на Северо-Востоке России, в определенном интервале интенсивности климатических стресс-факторов, в их тканях в 1,8–2,5 раза увеличивается общее содержание и, главное, более чем в 3 раза – структурное разнообразие биологически активных веществ (БАВ) регуляторного и защитного действия [Голдовский, 1941; Телятьев, 1987; Брехман, Нестеренко, 1988; Алексеев, 1994; Макаров, 1989]. Специальные исследования химического состава и питательной ценности кормовых трав Якутии [Егоров, 1954, 1969; Говоров, Торговкина, 1974; Макаров, 1989; Попова, 2003] хорошо согласуются с высказанными предположениями. Авторы, изучая преимущественно растения из семейства злаковых и осоковых, установили существование двух региональных кормовых профилей и двух типов растений: богатобелковых и бедноуглеводистых (сырого протеина до 15 %, безазотистых экстрактивных веществ меньше 40 %) и беднобелковых и высокоуглеводистых (сырого протеина менее 10 %, БЭВ около 50–60 %). Они сравнили свои данные с соответствующими данными других авторов, полученными в различных зонах европейской части СССР. В результате было установлено, что второй тип растений в сущности не отличается от злаков и осок в других областях страны по соотношению питательных веществ. Что касается первого типа, то он нигде ранее не был найден и, по-видимому, составляет региональную особенность растений Якутии. Около 65 % от общего числа проанализированных образцов злаковых и осоковых Якутии дали высокое и сверхвысокое содержание протеина и белка, тогда как этот показатель для лесолуговой зоны бывшей территории СССР составил 32 %, для лесостепной и степной зон – около 40 %, а для пустынной – 53 %.
В настоящее время можно считать твердо установленным, что отдельные группы кормовых растений Центральной Якутии (бобовые, злаки) отличаются более повышенным уровнем содержания протеина и белка и низким – клетчатки, по сравнению с растениями других районов России с менее континентальным климатом (Западная Сибирь, Дальний Восток и др.). Притом такая картина наблюдается соответственно во всех фазах роста и развития растений. Закономерное увеличение содержания белка и азотистых веществ при одновременном уменьшении клетчатки наблюдается и внутри самого региона по мере продвижения к северу [Макаров, 1989].
Обнаружена специфика и в биосинтезе углеводов. Установлено более высокое накопление растворимых углеводов и полисахаридов типа гемицеллюлоз в тканях аборигенных растений по сравнению с растениями аналогичных видов, произрастающих в средней полосе России [Егоров, 1954]. Как полагает автор, белки играют не только обменную, но и защитную роль, а более интенсивный биосинтез указанных форм углеводов связан с повышенной морозо- и зимостойкостью северных растений.
Специфика природных условий Якутии накладывает свой отпечаток на минеральный и микроэлементный состав местных растений. Уровень микроэлементов в отдельных растениях в значительной степени зависит от минерального состава воды и почвы. В этом отношении на территории Якутии выделяют три ландшафтные биогеохимические провинции. Это:
1) таежно-озерно-аласная, отличающаяся высоким содержанием натрия, калия, магния и хлора;
2) мелкодолинная заболоченная таежно-речная, очень бедная хлором и натрием;
3) крупнодолинная по поймам рек Лены, Вилюя, Амги и др., богатая кальцием и кремнекислотой; к ней же примыкают и отдельные таежные лугоболотные участки.
В отличие от растений других природных зон России злаковые, хвощовые и осоковые растения Якутии более обогащены железом, марганцем, медью, йодом, отчасти молибденом и цинком. Причем растения пониженных участков и замкнутых депрессий рельефа, как правило концентрируют значительно больше микроэлементов, чем растения повышенных и сухих мест обитания (лугостепи) [Егоров, 1969].
Таким образом, исследования химизма растений якутского региона отчетливо показали наличие у них заметных отклонений от широтно-зональных норм в синтезе и накоплении веществ основного обмена (белков, жиров, некоторых форм углеводов) в сторону повышения. Наряду с этим установлена специфика и в микроэлементном составе местных растений.
Выраженная региональная специфика отчетливо прослеживается также в отношении синтеза и накопления ФАВ. Так, алкалоидоносность является одним из существенных физиолого-биохимических свойств растений [Юнусов, 1948, 1974]. Согласно фактическому материалу отдельные виды растений якутской флоры синтезируют и накапливают очень много органических оснований, по процентному содержанию превышающих порою содержание алкалоидов в растениях более южных пунктов их ареалов [Макаров, 1989]. Полуколичественное изучение содержания алкалоидов и флавоноидов в 346 видах растений Якутии, проведенное А.А. Макаровым, выявило, что, например, по алкалоидоностности флора Центральной Якутии близка к флоре горных районов Казахстана и существенно превосходит флору Азербайджана и Средней Азии. Алкалоидоносными оказались и растения Крайнего Севера Якутии (бассейн Колымы, Яны). По мнению автора флора якутского региона может стать источником некоторых видов эффективного алкалоидоносного сырья.
Уже первые биохимические исследования около 70 видов лекарственных растений Якутии, проведенные в 1954 г. А.Д. Егоровым, показали высокое содержание в их листьях и других органах аскорбиновой кислоты (АК) и каротиноидов [Егоров, 1954]. Например, в спелых ягодах черной смородины содержание АК в условиях Центральной Якутии достигает 3,9 мг/г сырой массы, в Хакассии – до 2,0; в долине р. Енисей – до 2,5; в Западной Сибири – до 1,1–1,2 мг/г сырой массы [Рожко, 1966]. Причем, по данным А.Д. Егорова при продвижении растений на Крайний Север, т.е. повышении фактора экстремальности, содержание АК в их листьях еще увеличивается.
Вместе с тем, в рамках видоспецифичных пределов, прослеживается прямая зависимость между общим содержанием ФАВ (рис. 1), особенно их структурным разнообразием (рис. 2) и степенью экстремальности температурно-влажностных погодных условий в диапазоне величин kэкстр от 0,2 до 1,4.
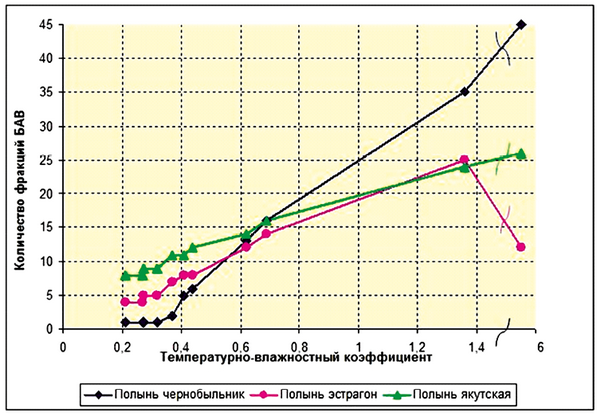
Рис. 1. Зависимость структурного разнообразия в надземных органах полыни от экстремальности метеорологических условий произрастания (температурно-влажностного коэффициента = t °C/влажность) в летние сезоны 1997–2008 гг.
Особого внимания заслуживает тот факт, что среди девяти групп ФАВ, обнаруженных во фракции эфирных масел, можно выделить такие, которые присутствуют хотя и видоспецифично, но почти во все годы исследования независимо от величины kэкстр погодных условий. Например, для полыни чернобыльник – это только производные камфоры. Для полыни эстрагон – производные бициклических непредельных и ароматических соединений, непредельные алифатические углеводороды и продукты их окисления. Производные пинена, моноциклических непредельных и ароматических соединений присутствуют во фракции эфирных масел полыни эстрагон во все годы кроме наиболее экстремального 2001 года. Для полыни «якутская» – производные камфоры, пинена, феландрена, бициклических непредельных и ароматических соединений, а также азулены и другие полимакроциклические соединения.
С другой стороны, – это фракции ФАВ, которые появляются только при высоких значениях kэкстр погодных условий (выше 0,4–0,5). Для полыни чернобыльник – это производные пинена, феландрена, бициклических непредельных и ароматических соединений, азулены и другие полимакроциклические соединения (при kэкстр > 0,5), непредельные алифатические углеводороды и продукты их окисления (при kэкстр > 0,6), производные моноциклических непредельных и ароматических соединений, дииновые и другие органические соединения (при kэкстр > 5). Для полыни эстрагон – дииновые соединения (при kэкстр > 0,4), производные феландрена (при kэкстр > 1,3), азулены (при 5 > kэкстр > 1) и прочие органические соединения (при kэкстр > 5). Для полыни «якутская» – непредельные алифатические углеводороды и продукты их окисления (при kэкстр>0,4), производные моноциклических непредельных и ароматических соединений (при 5 > kэкстр > 1) и прочие органические соединения (при kэкстр > 5). Причем первые из них составляют, как правило, значительную массовую долю всех ФАВ, а структурное разнообразие особенно вторых резко возрастает при повышении kэкстр погодных условий. Эти факты можно интерпретировать с позиций механизма сохранения способности организмов растений к самоорганизации (к адаптации и сохранению жизнеспособности) даже в условиях выраженной экстремальности среды. Как известно из теории самоорганизации систем [Пригожин, Стенгерс, 2003; Чернавский, 2004], для сохранения способности систем к самоорганизации в изменяющихся условиях среды в них должно возникать в особые периоды времени («периоды с обострением» [Князева, Курдюмов, 2002] состояние «динамического хаоса». Благодаря нему и становится возможным формирование при изменении условий среды новых адаптивных (диссипативных) состояний системы. По-видимому, вторая группа ФАВ во фракции эфирных масел полыней, наиболее гетерогенная по структуре, хотя и незначительная по общему количеству («минорная группа ФАВ») как раз и выполняет эту функцию формирования «динамического хаоса», обеспечивающего адаптацию организмов при резком увеличении степени экстремальности (неадекватности) среды. По-видимому, именно по этой причине увеличение структурного разнообразия ФАВ повышает общую устойчивость, обеспечивает выживание растений в условиях дефицита влаги [Кершенгольц и др., 2008]. Причем, сама величина интервала kэкстр, в котором сохраняется пропорциональность со структурным разнообразием ФАВ вторичного происхождения отражает, вероятно, величину адаптивного потенциала растений данного вида. При более высоких значениях kэкстр эти характеристики либо перестают расти (у полыни чернобыльника), либо снижаются (у полыни якутской, особенно у полыни эстрагона), что, вероятно, указывает на снижение их адаптивного потенциала (рис. 1 и 2).
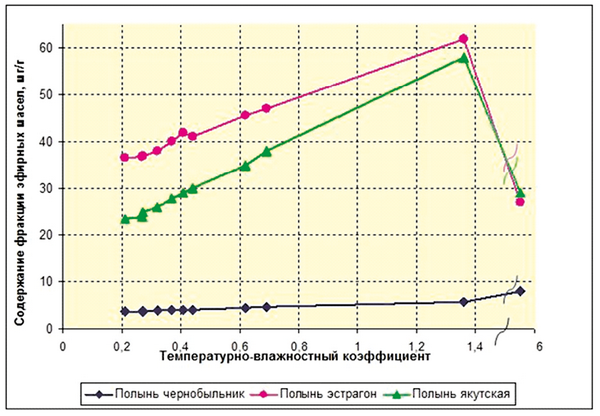
Рис. 2. Зависимость содержания эфирных масел в надземных органах полыни от экстремальности метеорологических условий произрастания (температурно-влажностного коэффициента = t °C/влажность) в летние сезоны 1997–2008 гг.
Также установлена степень и характер влияния сезонных изменений на накопление физиологически активных веществ в трех видах лишайников: Сladonia amaurocraea (Flk.) Shaer., Сladonia rangiferina (L.) Wed., Cladina stellaris (Opiz.) Brodo., произрастающих на территории Центральной Якутии. Определена активность пероксидазы, суммарное содержание низкомолекулярных антиоксидантов (НМАО), аскорбиновой кислоты (АК) и суммарное содержание полисахаридов.
Известно, что в процессе биохимической адаптации одними из ранних реакций клеток растений на снижение температуры окружающей среды являются адаптивные изменения состава жирных кислот в липидах и соотношения липидных фракций в мембранных структурах. К этим механизмам, прежде всего, относятся переориентировка метаболизма на усиленный биосинтез защитных соединений (антиоксидантов) и перестройка липидного компонента, связанная с увеличением синтеза полиненасыщенных жирных кислот, поддерживающих жидкокристаллическое состояние липидов мембран при более низких температурах. Как известно, в растениях Якутии одним из механизмов адаптации к экстремальным условиям является увеличение активности антиоксидантных систем [Кершенгольц, 1995; Кузьмина и др., 2001].
При адаптации к изменениям климатических факторов в растениях, в первую очередь, происходит повышение содержание НМАО. Было выявлено, что содержание НМАО у всех видов исследованных лишайников одинаково снижается с лета к осени: Сladonia amaurocraea на 25 %, Сladonia rangiferina на 8 % и Сladina stellaris на 51 % и постепенно повышается к зиме (рис. 3). Затем далее повышается к весне в среднем на 7 %. Это указывает на участие НМАО в процессах перестройки организма растений при подготовке к зимнему покою [Соловьева, Кузьмина, 2008].
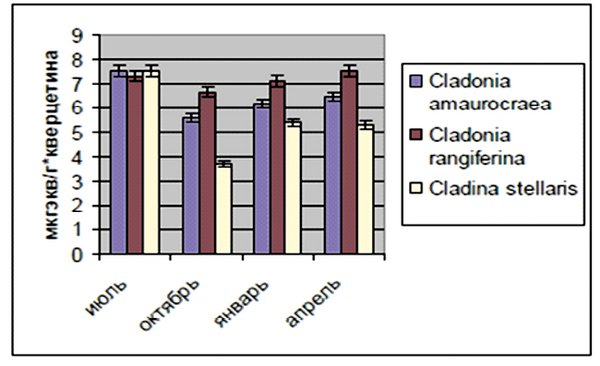
Рис. 3. Динамика накопления НМАО в слоевищах лишайников в зависимости от сезона года
Аскорбиновая кислота является важным компонентом биологической антиоксидантной системы, тесно взаимосвязанным с глутатионом (G-SH) и токоферолом. По литературным данным, содержание аскорбиновой кислоты в лишайниках р. Cladonia имеет примерно такую же динамику изменений в зависимости от сезона года (рис. 4), как и содержание НМАО [Соловьева, Кузьмина, 2008].
Пероксидаза является ферментом, также обладающим антиоксидантными свойствами. Это весьма динамичный фермент, чутко реагирующий при различных воздействиях на растительный организм. Большое значение придается пероксидазе в устойчивости растений к неблагоприятным факторам окружающей среды [Рубин и др., 1968]. По данным авторов [Соловьева, Кузьмина, 2008], активность пероксидазы в лишайниках Сladonia amaurocraea осенью снижается на 49 %, а у Сladonia rangiferina и Сladonia stellaris повышается на 13 и 7 % соответственно. С января по апрель у всех исследованных видов лишайников активность пероксидазы остается примерно на одном уровне.
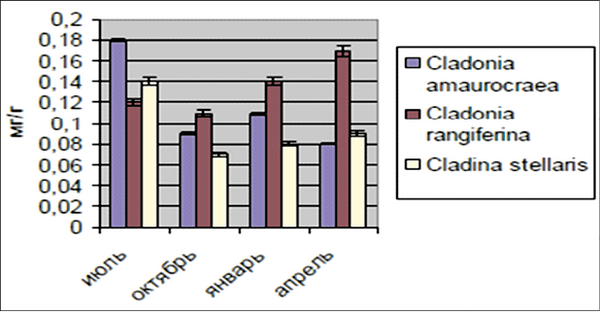
Рис. 4. Динамика активности аскорбиновой кислоты в слоевищах лишайников в зависимости от сезона года
Как известно, полисахариды также усиленно накапливаются при воздействии стрессовых факторов, так как они участвуют при стабилизации биомембран, образуя эффективный барьер от адгезии. Полисахариды имеют большое значение в строении мембраны клетки растений (в первую очередь, тилакоидных мембран), которые выполняют структурную и питательную функции. Они также способны связывать токсичные соединения, например, ионы тяжелых металлов. При использовании в медицине они обладают противоязвенными, ранозаживляющими свойствами, антибактериальным, антивирусным, противолучевым, противоопухолевым действием, поэтому изучение полисахаридов имеет большое практическое значение. У всех исследованных лишайников происходит накопление полисахаридов: у Сladonia amaurocraea – на 7 %, у Сladonia rangiferina – на 29 % и Сladina stellaris – на 10 %, а в зимний и весенний периоды – их постепенное расходование в среднем на 4,5 и 6 % соответственно (рис. 5) [Соловьева, Кузьмина, 2008.]
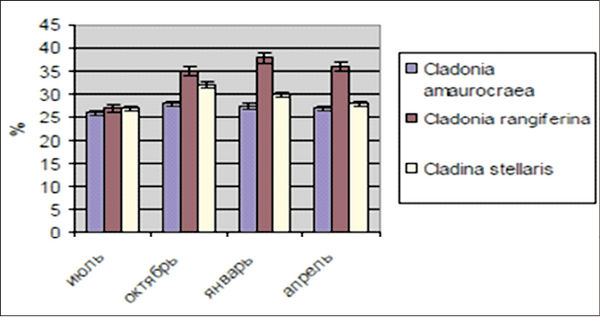
Рис. 5. Содержание полисахаридов в лишайниках в зависимости от сезона года
Накопление полисахаридов при стрессах связывают с их антиденатурационным действием на белково-липидные компоненты клеток, которые подвергаются дегидратации при стрессовых условиях. Результаты изменений по характеру и степени НМАО, пероксидазы, аскорбиновой кислоты и полисахаридов в годовом цикле роста и развития лишайников подтверждают защитную функцию данных биоантиоксидантов как эндогенного фактора обеспечения их зимостойкости. Таким образом, у лишайников одним из способов адаптации к сезонным изменениям (осень-зима) является увеличение содержания низкомолекулярных антиоксидантов, накопление и расходование полисахаридов.
Немаловажным является тот факт, что повышение устойчивости носит неспецифический характер в силу общности основных физиолого-биохимических механизмов адаптации к действию различных по природе стресс-факторов, т.е. развиваясь в ответ на действие одного экстремального фактора (например, климатического), такая адаптация приводит к повышению устойчивости данного организма и к действию других экстремальных факторов (например, радиационного или химиотоксического).
Поскольку эти ФАВ участвуют в регуляции биохимических реакций первичного обмена веществ (общего для различных видов организмов, включая человека) либо функционирования систем защищающих клетки и организм от действия экзо- или эндогенных токсинов (в том числе мутагенов), их относят к ФАВ неспецифического действия. Будучи выделенными из тканей одних видов растений (животных) в виде комплексов ФАВ, которые практически невозможно получить иным (химиосинтетическим, генноинженерным) путем, они способны проявлять свою регуляторную, защитную активность и по отношению к организмам других видов, включая человека [Гриневич и др., 1977; Кершенгольц и др., 2004]. Следует отметить, что некоторые ФАВ вообще образуются в процессе биотехнологической переработки биосырья.
Анализ литературных данных позволяет заключить, что синтез различных групп ФАВ активируется (или частично ингибируется) в зависимости от специфической природы стресс-фактора, взаимодополняя друг друга: при увеличении влажности и снижении температуры – флавоноидов; при увеличении температуры, но небольшом снижении влажности – аскорбиновой кислоты; при увеличении солнечной инсоляции (и небольшом снижении температуры) – рутина. При этом общий уровень антиоксидантной активности растет по мере повышения степени экстремальности условий произрастания, что и обеспечивает, в частности, повышение устойчивости организмов растений в этих условиях.